
หน้าที่ของการรุกรานของเยื่อหุ้มเซลล์ไซโตพลาสซึม เยื่อหุ้มเซลล์ไซโตพลาสซึม หน้าที่และโครงสร้างของมัน
เซลล์ – หน่วยโครงสร้างพื้นฐาน หน้าที่ และพันธุกรรมของการจัดระเบียบของสิ่งมีชีวิต ระบบสิ่งมีชีวิตเบื้องต้น เซลล์สามารถดำรงอยู่เป็นสิ่งมีชีวิตที่แยกจากกัน (แบคทีเรีย โปรโตซัว) หรือเป็นส่วนหนึ่งของเนื้อเยื่อของสิ่งมีชีวิตหลายเซลล์ คำว่า "เซลล์" ถูกเสนอโดยนักสำรวจชาวอังกฤษ Robert Hooke ในปี 1665
บทบัญญัติหลักของทฤษฎีเซลล์ถูกกำหนดโดยนักพฤกษศาสตร์ Schleiden (1838) และนักสัตววิทยา-สรีรวิทยา Schwann (1839) ในปี ค.ศ. 1858 Virchow ได้เสริมบทบัญญัติดังกล่าวด้วยข้อความเกี่ยวกับการแบ่งเซลล์
บทบัญญัติพื้นฐานของทฤษฎีเซลล์สมัยใหม่:
สิ่งมีชีวิตทั้งหมดประกอบด้วยเซลล์ เซลล์เป็นหน่วยของโครงสร้าง การทำงาน การสืบพันธุ์ และการพัฒนาสิ่งมีชีวิตส่วนบุคคล ไม่มีชีวิตนอกห้องขัง
เซลล์ของสิ่งมีชีวิตทุกชนิดมีความคล้ายคลึงกันในด้านโครงสร้างและองค์ประกอบทางเคมี
เซลล์สามารถเกิดขึ้นได้จากเซลล์ตามการแบ่งเท่านั้น
โครงสร้างเซลล์ของสิ่งมีชีวิตทั้งหมดเป็นหลักฐานที่แสดงถึงความเป็นเอกภาพของแหล่งกำเนิด
คำจำกัดความสมัยใหม่ของเซลล์:
เซลล์เป็นระบบชีววิทยาแบบเปิด ล้อมรอบด้วยเมมเบรนกึ่งซึมผ่านได้ ซึ่งประกอบด้วยนิวเคลียสและไซโตพลาสซึม มีความสามารถในการควบคุมตนเองและการสืบพันธุ์ด้วยตนเอง
สิ่งมีชีวิตสองประเภทมีโครงสร้างเซลล์ - โปรคาริโอต (แบคทีเรียและสาหร่ายสีน้ำเงินแกมเขียว) และยูคาริโอต (รูปที่ 2.1) เซลล์ยูคาริโอตประกอบด้วยอุปกรณ์พื้นผิว (เยื่อหุ้มเซลล์ไซโตพลาสซึม) ไซโตพลาสซึมและนิวเคลียส
เยื่อหุ้มเซลล์ไซโตพลาสซึม
เยื่อหุ้มเซลล์ไซโตพลาสซึมทำหน้าที่สำคัญหลายประการ: สิ่งกีดขวาง (การกำหนดขอบเขต), การควบคุม (ควบคุมการไหลของเมตาบอลิซึม), การขนส่ง (รับประกันการซึมผ่านแบบเลือกสรรของสารผ่านการขนส่งแบบพาสซีฟและแอคทีฟ), โครงสร้าง, เมแทบอลิซึม เยื่อชีวภาพประกอบด้วยไขมัน โปรตีน และ
คาร์โบไฮเดรต (รูปที่ 2.2) มีการเสนอแบบจำลองโครงสร้างของเยื่อหุ้มเซลล์ไซโตพลาสซึมหลายแบบจำลอง (แบบจำลอง "แซนวิช" - แบบจำลอง Danieli และ Dawson, แบบจำลอง Lenard เป็นต้น) เยื่อเมมเบรนมีหลายประเภทขึ้นอยู่กับการใช้งาน ในปัจจุบัน แบบจำลองฟลูอิด-โมเสกที่เสนอโดยซิงเกอร์-นิโคลสัน (1972) ถูกนำมาใช้เป็นพื้นฐาน ตามแบบจำลองนี้ เยื่อหุ้มประกอบด้วยชั้นไขมันสองโมเลกุล ซึ่งรวมถึงโมเลกุลโปรตีนด้วย
ไขมันเป็นสารที่ไม่ละลายน้ำ พวกมันมีหัวที่มีขั้ว (มีประจุ) และมีโซ่คาร์โบไฮเดรตที่ไม่มีประจุ (ไม่มีขั้ว) ยาว โมเลกุลของไขมันหันหน้าเข้าหากันโดยมีปลายที่ไม่มีขั้ว และขั้ว (หัว) ของพวกมันยังคงอยู่ด้านนอก ก่อตัวเป็นพื้นผิวที่ชอบน้ำ (รูปที่ 2.3)
โปรตีนเมมเบรนสามารถแบ่งออกเป็นสามกลุ่ม: อุปกรณ์ต่อพ่วง (เกี่ยวข้องอย่างอ่อนที่สุดกับเมมเบรน), จมอยู่ใต้น้ำ (กึ่งปริพันธ์) และซึมผ่าน (ปริพันธ์) ก่อตัวเป็นรูพรุนของเมมเบรน โปรตีนเมมเบรนแบ่งออกเป็นเอนไซม์ การขนส่ง โครงสร้าง และกฎระเบียบ
บนพื้นผิวด้านนอกของพลาสมาเมมเบรน โมเลกุลของโปรตีนและไขมันสัมพันธ์กับสายโซ่คาร์โบไฮเดรตเพื่อสร้างไกลโคคาลิกซ์ โซ่คาร์โบไฮเดรตทำหน้าที่เป็นตัวรับ เซลล์ได้รับความสามารถในการตอบสนองต่ออิทธิพลภายนอกโดยเฉพาะ ดังนั้นปฏิสัมพันธ์ของฮอร์โมนกับตัวรับ "ของมัน" จากภายนอกทำให้เกิดการเปลี่ยนแปลงโครงสร้างของโปรตีนอินทิกรัลซึ่งนำไปสู่การกระตุ้นการตอบสนองของเซลล์ โดยเฉพาะอย่างยิ่งการตอบสนองดังกล่าวสามารถแสดงออกมาในรูปแบบของ "ช่องทาง" ซึ่งสารละลายของสารบางชนิดเริ่มเข้าหรือออกจากเซลล์
หน้าที่ที่สำคัญอย่างหนึ่งของเมมเบรนคือเพื่อให้แน่ใจว่ามีการสัมผัสกันระหว่างเซลล์ภายในอวัยวะและเนื้อเยื่อ
ใต้พลาสมาเมมเบรนที่ด้านไซโตพลาสซึมจะมีชั้นเยื่อหุ้มสมองและโครงสร้างไฟบริลลาร์ในเซลล์ที่ให้ความเสถียรเชิงกลของเมมเบรน
ในเซลล์พืช นอกเมมเบรนมีโครงสร้างหนาแน่น - เยื่อหุ้มเซลล์ประกอบด้วยโพลีแซ็กคาไรด์ (เซลลูโลส)
คุณสมบัติที่สำคัญที่สุดอย่างหนึ่งของไซโตพลาสซึมเกี่ยวข้องกับความสามารถในการส่งผ่านสารต่าง ๆ เข้าหรือออกจากเซลล์ นี่เป็นสิ่งจำเป็นเพื่อรักษาความสอดคล้องขององค์ประกอบ โมเลกุลและไอออนขนาดเล็กผ่านเยื่อหุ้มเซลล์โดยการขนส่งแบบพาสซีฟและแอคทีฟ
การขนส่งแบบพาสซีฟเกิดขึ้นโดยไม่มีการใช้พลังงานผ่านการแพร่กระจาย การออสโมซิส และการแพร่กระจายแบบอำนวยความสะดวก (รูปที่ 2.4) การแพร่กระจาย –

การลำเลียงโมเลกุลและไอออนผ่านเมมเบรนจากบริเวณที่มีความเข้มข้นสูงไปยังบริเวณที่มีความเข้มข้นต่ำ ได้แก่ ตามแนวไล่ระดับความเข้มข้น หากสารละลายได้ในไขมันสูง สารเหล่านั้นจะทะลุเซลล์โดยการแพร่กระจายอย่างง่าย (ออกซิเจน คาร์บอนไดออกไซด์) การแพร่กระจายของน้ำผ่านเยื่อกึ่งซึมผ่านเรียกว่าออสโมซิส น้ำยังสามารถผ่านรูเมมเบรนที่เกิดจากโปรตีนและขนส่งโมเลกุลของไอออนและสารที่ละลายอยู่ในนั้น การแพร่กระจายที่อำนวยความสะดวกคือการขนส่งสารที่ไม่ละลายในไขมันและไม่ผ่านรูขุมขนผ่านช่องไอออนโดยใช้โปรตีนตัวพา
การขนส่งสารอย่างแข็งขันผ่านเมมเบรนเกิดขึ้นจากการใช้พลังงาน ATP และการมีส่วนร่วมของโปรตีนพาหะ มันดำเนินการกับการไล่ระดับความเข้มข้น (นี่คือวิธีการขนส่งกรดอะมิโน, น้ำตาล, โพแทสเซียม, โซเดียม, แคลเซียมไอออน ฯลฯ ) ตัวอย่างของการขนส่งแบบแอคทีฟคือการทำงานของปั๊มโพแทสเซียมโซเดียม ความเข้มข้นของ K ภายในเซลล์สูงกว่าภายนอก 10-20 เท่าและ Na - ในทางกลับกัน เพื่อรักษาความเข้มข้นนี้ Na ไอออนสามตัวจะถูกถ่ายโอนจากเซลล์ทุกๆ สอง K ไอออนเข้าสู่เซลล์ กระบวนการนี้เกี่ยวข้องกับโปรตีนในเมมเบรน ซึ่งทำหน้าที่เป็นเอนไซม์ที่สลาย ATP และปล่อยพลังงานที่จำเป็นสำหรับปั๊มทำงาน (รูปที่ 2.5. A)
การถ่ายโอนโมเลกุลขนาดใหญ่และอนุภาคขนาดใหญ่เข้าไปในเซลล์นั้นดำเนินการโดยเอ็นโดไซโทซิส และการกำจัดออกจากเซลล์โดยเอ็กโซไซโทซิส
ในระหว่างกระบวนการเอนโดโทซิส (รูปที่ 2.5 B) เมมเบรนจะก่อให้เกิดการรุกรานหรือการเจริญเติบโตของเซลล์ ซึ่งจากนั้นจะรวมตัวกันและเปลี่ยนเป็นถุงภายในเซลล์ที่มีผลิตภัณฑ์ที่เซลล์จับไว้ กระบวนการนี้เกิดขึ้นพร้อมกับการใช้พลังงาน ATP endocytosis มีสองประเภท - phagocytosis (การดูดซึมของอนุภาคขนาดใหญ่โดยเซลล์) และ pinocytosis (การดูดซึมสารของเหลว)
เมมเบรนมีส่วนร่วมในการกำจัดสารออกจากเซลล์ในระหว่างกระบวนการเอ็กโซไซโทซิส ด้วยวิธีนี้ ฮอร์โมน โปรตีน หยดไขมัน ฯลฯ จะถูกกำจัดออกจากเซลล์
ใต้ผนังเซลล์ของแบคทีเรียคือเยื่อหุ้มเซลล์ไซโตพลาสซึม (CPM) แยกเนื้อหาของเซลล์ออกจากผนังเซลล์และเป็นโครงสร้างสำคัญของเซลล์
ความหนาของ CPM ของแบคทีเรียมักจะอยู่ที่ประมาณ 6-8 นาโนเมตร คิดเป็นสัดส่วนมากถึง 15% ของมวลแห้งของเซลล์ ประกอบด้วยไขมัน (15-45%) โปรตีน (45-60%) และคาร์โบไฮเดรตจำนวนเล็กน้อย (ประมาณ 10%) ไขมันจะแสดงโดยฟอสโฟลิปิด - มากถึง 30% ของน้ำหนักแห้งของเมมเบรน ในหมู่พวกเขา phosphatidyl glycerol และ diphosphatidyl glyceride (cardiolipin) มีอำนาจเหนือกว่าซึ่งเป็นองค์ประกอบสำคัญของเยื่อหุ้มไมโตคอนเดรียของยูคาริโอต ปริมาณที่น้อยกว่าประกอบด้วยฟอสฟาติดิลโนซิทอลและฟอสฟาไทด์อิล-
เอทานอลเอมีน นอกจากฟอสโฟลิปิดแล้ว ยังพบไกลโคลิปิดหลายชนิด แคโรทีนอยด์ และควิโนนจำนวนเล็กน้อยในเยื่อหุ้มเซลล์ ในองค์ประกอบของไขมันที่ได้จากกลีเซอรอลพบว่ากรดไขมันผิดปกติสำหรับเยื่อหุ้มเซลล์ - อิ่มตัวหรือไม่อิ่มตัวเชิงเดี่ยวด้วยอะตอมคาร์บอน 16-18 อะตอมรวมถึงกรดที่ไม่พบในเยื่อหุ้มยูคาริโอต - ไซโคลโพรเพนและกรดไขมันกิ่งก้านที่มีอะตอมคาร์บอน 15-17 อะตอม ชุดของกรดไขมันตลอดจนไขมันเมมเบรนที่ประกอบด้วยกรดไขมันเหล่านี้เป็นสายพันธุ์เฉพาะสำหรับโปรคาริโอต
ไขมันเมมเบรนเป็นโมเลกุลขั้วโลกขนาดเล็กที่มีกลุ่มที่ชอบน้ำ (หัว) และกลุ่มที่ไม่ชอบน้ำ (หาง) ในสภาพแวดล้อมที่เป็นน้ำพวกมันจะก่อตัวเป็นชั้น bimolecular แบบปิดตามธรรมชาติ - เป็นชั้นสอง ชั้นนี้ทำหน้าที่เป็นอุปสรรคสำคัญต่อไอออนและสารประกอบเชิงขั้ว ลิพิดที่จัดเป็นชั้นสองโมเลกุลจะสร้างพื้นฐานโครงสร้างของเมมเบรน รักษาเสถียรภาพทางกล และให้ความไม่ชอบน้ำแก่มัน
โปรตีนประกอบขึ้นเป็นมากกว่าครึ่งหนึ่งของมวลแห้งของเมมเบรน มีมากกว่า 20 ประเภทที่แตกต่างกัน ขึ้นอยู่กับความแตกต่างในความแข็งแรงของพันธะกับไขมันและตำแหน่งในเมมเบรน โปรตีนจะถูกแบ่งออกเป็นอินทิกรัลและอุปกรณ์ต่อพ่วง โปรตีนอินทิกรัลถูกแช่อยู่ในบริเวณที่ไม่ชอบน้ำของเมมเบรนซึ่งพวกมันสร้างพันธะมากมายกับสายโซ่ไฮโดรคาร์บอนของไขมัน
สร้างคอมเพล็กซ์ไลโปโปรตีน โปรตีนส่วนปลายจะถูกแปลเป็นภาษาท้องถิ่นบนพื้นผิวของชั้นที่ชอบน้ำและมักจะเกาะติดกับโปรตีนอินทิกรัล (รูปที่ 3.14)
รูปที่.3.14. โครงสร้างของเยื่อหุ้มเซลล์ไซโตพลาสซึม: 1 - ไขมัน; 2 - ไกลโคโปรตีน; 3 - โปรตีนส่วนปลาย; 4 - โปรตีนอินทิกรัล
ขึ้นอยู่กับหน้าที่ของมันภายในเมมเบรน โปรตีนของเมมเบรนสามารถแบ่งออกเป็นสองกลุ่ม: แบบโครงสร้างและไดนามิก
หน้าที่ของโปรตีนโครงสร้างจำกัดอยู่ที่การรักษาความสมบูรณ์ของโครงสร้างของเมมเบรน ตั้งอยู่บนพื้นผิวของชั้นไขมันที่ชอบน้ำซึ่งทำหน้าที่เป็นผ้าพันแผลระดับโมเลกุล
ไดนามิกโปรตีนรวมถึงโปรตีนที่เกี่ยวข้องโดยตรงกับกระบวนการทั้งหมดที่เกิดขึ้นบนเมมเบรน แบ่งออกเป็นสามประเภท ได้แก่ การขนส่ง เกี่ยวข้องกับการขนส่งสารประกอบเข้าและออกจากเซลล์ ตัวเร่งปฏิกิริยาซึ่งทำหน้าที่ของเอนไซม์ในปฏิกิริยาที่เกิดขึ้นบนเมมเบรน โปรตีนตัวรับที่จับกับสารประกอบบางชนิดโดยเฉพาะ (สารพิษ ฮอร์โมน) ที่ด้านนอกของเมมเบรน
คาร์โบไฮเดรตในเมมเบรนไม่อยู่ในสถานะอิสระ แต่เชื่อมโยงกับโปรตีนและไขมันเป็นไกลโคโปรตีน พวกเขาเป็นเหมือน
ตามกฎแล้ว พวกมันจะถูกแปลเป็นภาษาท้องถิ่นเฉพาะบนพื้นผิวด้านนอกของเมมเบรน และทำหน้าที่เป็นตัวรับการรับรู้สำหรับปัจจัยด้านสิ่งแวดล้อม
เมมเบรนไซโตพลาสซึมของแบคทีเรีย เช่นเดียวกับเยื่อหุ้มชีวภาพอื่นๆ คือโครงสร้างผลึกเหลวที่ไม่สมมาตร ความไม่สมดุลเกิดจากความแตกต่างในโครงสร้างทางเคมีของโมเลกุลโปรตีนและตำแหน่งของมันในชั้นไขมันของเมมเบรน โปรตีนบางชนิดอยู่บนพื้นผิวของชั้นสองชั้น ส่วนโปรตีนบางชนิดจะถูกแช่อยู่ในความหนาของมัน และบางชนิดก็ผ่านจากด้านในไปยังพื้นผิวด้านนอกของชั้นสองชั้น ในทางกลับกัน การวางแนวที่กำหนดไว้อย่างเคร่งครัดของโปรตีนเมมเบรนนั้นเกิดจากการสังเคราะห์และรวมเข้ากับเมมเบรนแบบไม่สมมาตร พื้นผิวด้านนอกและด้านในของเมมเบรนก็มีกิจกรรมของเอนไซม์ต่างกันเช่นกัน ขึ้นอยู่กับเงื่อนไข (เช่น อุณหภูมิ) CPM อาจอยู่ในสถานะเฟสที่แตกต่างกัน: กลายเป็นของเหลวหรือเป็นผลึก ในระหว่างการเปลี่ยนเฟสผลึกเหลวหนึ่งไปอีกเฟสหนึ่ง การเคลื่อนตัวของส่วนประกอบเมมเบรนและความหนาแน่นของการบรรจุจะเปลี่ยนไป ซึ่งในทางกลับกัน จะนำไปสู่การหยุดชะงักในกิจกรรมการทำงานของส่วนประกอบนั้น
โครงสร้างและหน้าที่ของเมมเบรนไซโตพลาสซึม เพื่ออธิบายธรรมชาติและกลไกของการทำงานหลายอย่างของ CPM สิ่งที่เหมาะสมที่สุดคือแบบจำลองของเหลว-โมเสคของการจัดระเบียบของเยื่อหุ้มชีวภาพ เสนอโดย R. Singer และ A. Nicholson ในปี 1972 ตามแบบจำลองนี้ เมมเบรนแบ่งออกเป็นสองส่วน - สารละลายเชิงมิติของโปรตีนและไขมันทรงกลมที่มุ่งเน้นในลักษณะใดลักษณะหนึ่ง ไขมันก่อตัวเป็นชั้นสองชั้น โดยที่ “หัว” ของโมเลกุลที่ชอบน้ำหันหน้าออกด้านนอก และ “หาง” ที่ไม่ชอบน้ำจะถูกจุ่มลงในความหนาของเมมเบรน ในขณะที่มีความยืดหยุ่นเพียงพอ ไขมันของเมมเบรนและโปรตีนหลายชนิดเคลื่อนที่อย่างอิสระในชั้นสองชั้น แต่จะเคลื่อนที่ไปในทิศทางด้านข้างเท่านั้น (การแพร่กระจายด้านข้าง) ในทิศทางตามขวาง เช่น จากพื้นผิวด้านหนึ่งของเมมเบรนไปอีกด้านหนึ่ง โปรตีนไม่สามารถเคลื่อนที่ได้ และไขมันจะเคลื่อนที่ช้ามาก (ทุกๆ สองสามชั่วโมง) สาเหตุของการไม่มีหรือกิจกรรมต่ำของการแพร่กระจายตามขวางดูเหมือนจะเกิดจากการกระจายตัวของไขมันแบบไม่สมมาตร:
ไขมันบางชนิดมีมากที่ชั้นนอกของชั้นสองชั้น ในขณะที่บางชนิดมีมากที่ชั้นใน ผลที่ตามมาคือความหนาแน่นของอิเล็กตรอนไม่เท่ากัน (การนำไฟฟ้า) ของชั้นสองชั้นในทิศทางตามขวาง
CPM อยู่ในสถานะผลึกเหลวหรือสถานะเป็นของเหลวภายใต้สภาวะบางอย่างที่เรียกว่าเท่านั้น
อุณหภูมิทางชีวภาพ เมื่ออุณหภูมิลดลง (ต่ำกว่าจุดหลอมเหลว Tm) ไขมันจะเปลี่ยนเป็นสถานะผลึก ระดับความหนืดจะเพิ่มขึ้นจนกระทั่งเมมเบรนแข็งตัว อุณหภูมิที่ทำให้เมมเบรนแข็งตัวจะถูกกำหนดโดยเนื้อหาที่ไม่อิ่มตัวและ
กรดไขมันกิ่งก้าน ยิ่งมีอยู่ในเมมเบรนมากเท่าไร อุณหภูมิการเปลี่ยนผ่านของไขมันจากสถานะผลึกเหลวไปเป็นสถานะผลึกก็จะยิ่งต่ำลงเท่านั้น
โปรคาริโอตมีความสามารถในการควบคุมการไหลของเมมเบรนโดยการเปลี่ยนจำนวนพันธะคู่และความยาวสายโซ่ของโมเลกุลกรดไขมัน ดังนั้น ในเชื้อ E. coli เมื่ออุณหภูมิสิ่งแวดล้อมลดลงจาก 42° C เป็น 27° C อัตราส่วนของกรดไขมันอิ่มตัวและไม่อิ่มตัวในเมมเบรนจะลดลงจาก 1.6 เป็น 1.0 กล่าวคือ ปริมาณกรดไขมันไม่อิ่มตัวจะถึงระดับที่ คนที่อิ่มตัว สิ่งนี้จะช่วยป้องกันการเพิ่มขึ้นของความหนืดและทำให้แน่ใจว่าเซลล์คงกิจกรรมทางสรีรวิทยาไว้ที่อุณหภูมิต่ำ
CPM ทำหน้าที่สำคัญมากมายในโปรคาริโอต ส่วนใหญ่จะถูกกำหนดโดยโปรตีนที่มีการแปลซึ่งทำหน้าที่เป็นช่องทางตัวรับตัวสร้างพลังงานใหม่เอนไซม์ฟังก์ชั่นการขนส่งและอื่น ๆ CPM เป็นตัวกั้นการดูดซึมหลัก ซึ่งเนื่องจากมีกลไกการขนส่งเมมเบรน จึงคัดเลือกขนส่งสารเข้าสู่เซลล์และกำจัดผลิตภัณฑ์เมตาบอลิซึมออกจากเซลล์ ความสามารถในการซึมผ่านแบบเลือกสรรของ CPM เกิดจากการซึมผ่านของสารตั้งต้นที่จำเพาะอยู่ในนั้น ซึ่งขนส่งสารอินทรีย์และแร่ธาตุต่างๆ ผ่านเมมเบรนอย่างแข็งขัน CPM ประกอบด้วยเอนไซม์สำหรับการสังเคราะห์ทางชีวภาพของไขมันในเยื่อหุ้มเซลล์และโมเลกุลขนาดใหญ่ที่ประกอบเป็นผนังเซลล์ เยื่อหุ้มชั้นนอก และแคปซูล CPM เป็นที่ตั้งของเอนไซม์รีดอกซ์ที่ดำเนินการ
การขนส่งอิเล็กตรอน ฟอสโฟรีเลชันแบบออกซิเดชันและสังเคราะห์แสง การสร้างพลังงานไฟฟ้าเคมี
ศักย์ของเมมเบรน (A// +) และสารเคมี (ATP) CPM
n
ทำหน้าที่สำคัญในการสังเคราะห์ทางชีวภาพและการเคลื่อนย้ายโปรตีนที่ถูกหลั่งโดยแบคทีเรียแกรมลบ การสังเคราะห์ทางชีวภาพของโปรตีนเหล่านี้ดำเนินการกับไรโบโซมที่ติดอยู่กับ CPM แบคทีเรียแกรมลบมีโปรตีนตัวรับพิเศษบน CPM ที่ "รับรู้" สัญญาณจากหน่วยย่อยไรโบโซมขนาดใหญ่เกี่ยวกับการเกาะติดของไรโบโซมและจุดเริ่มต้นของการสังเคราะห์โปรตีน โปรตีนตัวรับเมมเบรนมีปฏิกิริยากับหน่วยย่อยขนาดใหญ่ของไรโบโซมซึ่งเกิดคอมเพล็กซ์ไรโบโซมเมมเบรนขึ้นซึ่งการสังเคราะห์โปรตีนที่หลั่งออกมาจะดำเนินการ ด้วยวิธีนี้ E. coli สังเคราะห์อัลคาไลน์ฟอสฟาเตส Bac subtilis - อะไมเลส CPM ยังช่วยให้แน่ใจว่ามีการถ่ายโอนโปรตีนเหล่านี้ไปยังปริพลาสมิกสเปซ CPM มีบทบาทสำคัญในการควบคุมการแบ่งเซลล์ โครโมโซมและการจำลองพลาสมิด และการแยกองค์ประกอบทางพันธุกรรมเหล่านี้ระหว่างเซลล์ลูกสาวที่สร้างขึ้นใหม่ในเวลาต่อมา
โปรคาริโอตทั้งหมดพร้อมกับเยื่อหุ้มเซลล์ไซโตพลาสซึมมีอนุพันธ์ - เยื่อหุ้มเซลล์ที่ทำหน้าที่พิเศษ เมมเบรนไซโตพลาสซึมสามารถสร้างการบุกรุกได้ทุกประเภท (การบุกรุก) การรุกรานเหล่านี้ประกอบขึ้นเป็นเยื่อหุ้มเซลล์ซึ่งมีความยาว บรรจุภัณฑ์ และตำแหน่งที่แตกต่างกันในไซโตพลาสซึม พวกเขาสามารถรวบรวมเป็นลูกบอลที่ซับซ้อน - ก่อตัวเป็นลาเมลลาร์, รังผึ้งหรือท่อ เมมเบรนที่ซับซ้อนน้อยกว่าจะอยู่ในรูปของลูปหรือท่อธรรมดาที่มีความยาวต่างกัน โดยไม่คำนึงถึงความซับซ้อนของการจัดระเบียบของเยื่อหุ้มเซลล์ในเซลล์พวกมันล้วนเป็นอนุพันธ์ของเยื่อหุ้มเซลล์ไซโตพลาสซึม ขนาดของพื้นผิวที่ใช้งานอยู่นั้นเกินกว่าเยื่อหุ้มเซลล์ไซโตพลาสซึม นี่เป็นเหตุให้ตัดสินกิจกรรมการทำงานที่ยอดเยี่ยมของโครงสร้างเหล่านี้ในเซลล์
อุปกรณ์เมมเบรนในเซลล์ที่อุดมสมบูรณ์เป็นพิเศษพบได้ในแบคทีเรียที่ตรึงไนโตรเจนและสังเคราะห์แสง บรูเซลลา และแบคทีเรียไนตริไฟอิง แบคทีเรียสังเคราะห์แสง (Rhodospirillum rubrum) มีเยื่อหุ้มที่มีลักษณะคล้ายฟองอากาศปิด - ถุง การก่อตัวของพวกมันเริ่มต้นด้วยการรุกรานของเยื่อหุ้มเซลล์ไซโตพลาสซึมซึ่งต่อมาก่อตัวเป็นหลอด มีการบีบรัดปรากฏบนท่อโดยแบ่งเป็นฟองอากาศหลายชุด ถุงเหล่านี้เรียกว่าโครมาโตฟอร์ ประกอบด้วยเม็ดสีที่ดูดซับแสง - แบคทีเรียคลอโรฟิลล์และแคโรทีนอยด์, เอนไซม์ขนส่งอิเล็กตรอน - ยูบิควิโนนและไซโตโครม, ส่วนประกอบของระบบฟอสโฟรีเลชั่น ในโปรคาริโอตสังเคราะห์แสงบางชนิดโดยเฉพาะอย่างยิ่งในแบคทีเรียกำมะถันสีม่วงและไซยาโนแบคทีเรียเครื่องมือสังเคราะห์แสงจะถูกแสดงโดยกองของเยื่อหุ้มที่มีรูปร่างแบนและโดยการเปรียบเทียบกับแกรนาของคลอโรพลาสต์ของพืชสีเขียวเรียกว่าไทลาคอยด์ (รูปที่ 3.15)  โดยให้ความเข้มข้นของเม็ดสีสังเคราะห์แสง เอนไซม์ของห่วงโซ่การขนส่งอิเล็กตรอน และระบบฟอสโฟรีเลชั่น คุณสมบัติของไซยาโนแบคทีเรียไทลาคอยด์คือขาดการเชื่อมต่อกับเยื่อหุ้มเซลล์ไซโตพลาสซึม นี่เป็นโปรคาริโอตกลุ่มเดียวที่มีระบบเมมเบรนที่แตกต่าง
โดยให้ความเข้มข้นของเม็ดสีสังเคราะห์แสง เอนไซม์ของห่วงโซ่การขนส่งอิเล็กตรอน และระบบฟอสโฟรีเลชั่น คุณสมบัติของไซยาโนแบคทีเรียไทลาคอยด์คือขาดการเชื่อมต่อกับเยื่อหุ้มเซลล์ไซโตพลาสซึม นี่เป็นโปรคาริโอตกลุ่มเดียวที่มีระบบเมมเบรนที่แตกต่าง
ในแบคทีเรียไนตริไฟติ้งอุปกรณ์เมมเบรนในเซลล์มีรูปแบบของแผ่นหรือแผ่นซึ่งประกอบด้วยถุงแบน (รูปที่ 3.16)  ในบรรดาเยื่อหุ้มเซลล์ในเซลล์ เมโซโซมมีโครงสร้างที่ซับซ้อนที่สุด มีลักษณะเป็นท่อบิดเป็นเกลียว แบนหรือเป็นทรงกลม Mesosomes เกิดขึ้นระหว่างการแบ่งเซลล์ในบริเวณที่เกิดกะบังตามขวาง พวกมันมีส่วนร่วมในการจำลองโครโมโซมและการกระจายจีโนมระหว่างเซลล์ลูก และในการสังเคราะห์สารผนังเซลล์ เพื่อเข้าร่วม
ในบรรดาเยื่อหุ้มเซลล์ในเซลล์ เมโซโซมมีโครงสร้างที่ซับซ้อนที่สุด มีลักษณะเป็นท่อบิดเป็นเกลียว แบนหรือเป็นทรงกลม Mesosomes เกิดขึ้นระหว่างการแบ่งเซลล์ในบริเวณที่เกิดกะบังตามขวาง พวกมันมีส่วนร่วมในการจำลองโครโมโซมและการกระจายจีโนมระหว่างเซลล์ลูก และในการสังเคราะห์สารผนังเซลล์ เพื่อเข้าร่วม
มีโซโซมในการแบ่งเซลล์ถูกระบุโดยการเชื่อมต่อกับ DNA ของนิวเคลียส เมโซโซมที่ได้รับการพัฒนาอย่างดีจะพบได้ในแบคทีเรียแกรมบวกเท่านั้น
ข้อมูลที่สะสมจนถึงปัจจุบันแสดงให้เห็นว่าโครงสร้างเมมเบรนของแบคทีเรียมีความแตกต่างเพียงพอและรับประกันกระบวนการเผาผลาญต่างๆ ในเซลล์
- ไซโตพลาสซึมและการรวมไซโตพลาสซึม
ในไซโตพลาสซึมจะแยกเศษส่วนออกเป็นสองส่วน หนึ่งในนั้นคือองค์ประกอบโครงสร้าง: ไรโบโซม แอโรโซม
คาร์บอกซีโซม การรวมการจัดเก็บ อุปกรณ์ทางพันธุกรรม ส่วนอีกส่วนหนึ่งประกอบด้วยส่วนผสมที่ซับซ้อนของ RNA ที่ละลายน้ำได้ โปรตีนของเอนไซม์ เม็ดสี แร่ธาตุ ผลิตภัณฑ์และสารตั้งต้นของปฏิกิริยาเมตาบอลิซึม เศษส่วนนี้เรียกว่าไซโตซอล
เนื่องจากมีสารประกอบอินทรีย์หลายชนิดไซโตพลาสซึมของเซลล์แบคทีเรียจึงมีความหนืดเพิ่มขึ้น มากกว่าความหนืดของน้ำ 800-8,000 เท่า (เข้าใกล้ความหนืดของกลีเซอรีน) เซลล์อายุน้อยที่อยู่ในระยะแล็กหรือระยะเริ่มต้นของระยะลอการิทึมมีความหนืดของไซโตพลาสซึมต่ำกว่า ในผู้สูงอายุ ความหนืดจะเพิ่มขึ้น มีลักษณะคล้ายเจลที่มีความคงตัว ระดับความหนืดของไซโตพลาสซึมไม่เพียงบ่งบอกถึงอายุของเซลล์เท่านั้น แต่ยังรวมถึงกิจกรรมทางสรีรวิทยาของมันด้วย การเพิ่มขึ้นของความหนืดของไซโตพลาสซึมในวัฒนธรรมเก่าเป็นปัจจัยหนึ่งที่ทำให้กิจกรรมทางสรีรวิทยาของเซลล์ลดลง ไซโตพลาสซึมเป็นสื่อที่เชื่อมต่อโครงสร้างภายในเซลล์ทั้งหมดให้เป็นระบบเดียว
ไรโบโซม ไซโตพลาสซึมของเซลล์แบคทีเรียมีโครงสร้างทรงกลมอยู่ตลอดเวลา ขนาด 15-20 นาโนเมตร โดยมีน้ำหนักโมเลกุล 3106
ไรโบโซมประกอบด้วยไรโบโซมอาร์เอ็นเอ 60-65% และโปรตีน 35-40% หลังอุดมไปด้วยกรดอะมิโนพื้นฐาน ในระหว่างการปั่นแยกด้วยความเข้มข้นสูง ไรโบโซมของแบคทีเรียจะตกลงตัวในอัตราประมาณ 70 หน่วย Svedberg (S)7 ซึ่งเป็นสาเหตุว่าทำไมพวกมันจึงถูกเรียกว่า 708-ไรโบโซม ไรโบโซมไซโตพลาสซึมของยูคาริโอตมีขนาดใหญ่กว่าและเรียกว่าไรโบโซม 80S (ค่าคงที่การตกตะกอนของพวกมันคือ 80S)
ไรโบโซมแต่ละอันประกอบด้วยสองหน่วยย่อย: 30S และ 50S ซึ่งมีขนาดโมเลกุล RNA และปริมาณโปรตีนที่แตกต่างกัน หน่วยย่อยขนาดใหญ่ (50S) ประกอบด้วยโมเลกุล rRNA สองโมเลกุล - 5S และ 23S และ 35 โมเลกุลของโปรตีนต่างๆ หน่วยย่อยขนาดเล็ก (30S) ประกอบด้วยหนึ่งโมเลกุลที่มี 16 rRNA และ 21 โมเลกุลของโปรตีนประเภทต่างๆ จำนวนไรโบโซมในเซลล์ไม่คงที่ - ตั้งแต่ 5,000 ถึง 90,000 ขึ้นอยู่กับอายุของเซลล์และเงื่อนไขของการเพาะเลี้ยงแบคทีเรีย จำนวนเงินขั้นต่ำจะอยู่ที่จุดเริ่มต้นของระยะล่าช้าและสูงสุด - ในระยะเอ็กซ์โพเนนเชียลของการเติบโตของวัฒนธรรม ในเชื้อ E. coli ในช่วงระยะเวลาของการเจริญเติบโตบนอาหารเลี้ยงเชื้อที่สมบูรณ์นั้น ไรโบโซม 5-6 ตัวจะถูกสังเคราะห์ใน 1 วินาที ส่วนใหญ่ในไซโตพลาสซึมของแบคทีเรียจะอยู่ในสถานะอิสระและที่เหลือก็คือ
S = 1 หน่วย swedberg = หน่วยสนาม 10"13 ซม.
รวมตัวกันด้วยสาย Messenger RNA ให้เป็นโพลีโซม จำนวนไรโบโซมในโพลีโซมสามารถเข้าถึงได้หลายโหล สิ่งนี้บ่งชี้ถึงกิจกรรมการสังเคราะห์โปรตีนที่สูงของเซลล์ เนื่องจากไรโบโซมเป็นที่ตั้งของการสังเคราะห์โปรตีน มีการเปรียบเทียบว่าโปรตีนเหล่านี้เป็น “โรงงาน”.
แก๊สแวคิวโอล (แอโรโซม) โครงสร้างเหล่านี้เป็นลักษณะเฉพาะของแบคทีเรียในน้ำและในดินบางชนิดเท่านั้น พบได้ในแบคทีเรียโฟโตโทรฟิกซัลเฟอร์ แบคทีเรียใยไม่มีสี และแบคทีเรียในสกุล Renobacter ในเซลล์มีมากถึง 40-60 ตัว (รูปที่ 3.17) แวคิวโอลของแก๊สถูกล้อมรอบด้วยบาง 
ข้าว. 3.17. เซลล์ Renobacter vocuolatum มีแอโรโซม (กำลังขยาย x 70,000)
เมมเบรนโปรตีน ประกอบด้วยฟองก๊าซซึ่งมีจำนวนไม่คงที่ องค์ประกอบและความดันของก๊าซในฟองอากาศและแอโรโซมโดยทั่วไปจะพิจารณาจากปริมาณของก๊าซที่ละลายในสิ่งแวดล้อม แอโรโซมอยู่ในสถานะอัดหรือเต็มไปด้วยตัวกลางก๊าซ สภาพของพวกเขาถูกควบคุมโดยความดันอุทกสถิตของสิ่งแวดล้อม ความดันที่เพิ่มขึ้นอย่างรวดเร็วทำให้เกิดการบีบอัดของแอโรโซมและเซลล์สูญเสียการลอยตัว
แอโรโซมควบคุมการลอยตัวของเซลล์ ทำให้สามารถเคลื่อนเซลล์ไปยังสภาวะการเติมอากาศ แสงสว่าง และสารอาหารที่เอื้ออำนวย คุณสมบัติพิเศษคือการทำงานเพียงครั้งเดียวเมื่อเติมแก๊ส หลังจากการบีบอัดภายใต้อิทธิพลของแรงดันอุทกสถิตพวกเขาจะไม่ถูกเติมด้วยแก๊สและ
จะค่อยๆถูกทำลายไป เซลล์สามารถสืบพันธุ์ได้โดยการสร้างใหม่เท่านั้น
เมื่อแอโรโซมเต็มไปด้วยก๊าซ แบคทีเรียจะถูกกักไว้บนผิวน้ำ เมื่อพวกมันถูกบีบอัด พวกมันจะจมลงในความหนาหรือตกลงไปที่ด้านล่างของอ่างเก็บน้ำ วิธีการเคลื่อนไหวที่เป็นเอกลักษณ์นี้ได้รับการพัฒนาในกระบวนการวิวัฒนาการโดยส่วนใหญ่อยู่ในแบคทีเรียที่ไม่มีแฟลเจลลา และด้วยเหตุนี้จึงมีความสามารถในการเคลื่อนไหวอย่างแข็งขัน
ไฟโคบิลิโซม โครงสร้างภายในเซลล์เหล่านี้เป็นลักษณะของไซยาโนแบคทีเรีย พวกมันมีรูปแบบของเม็ดที่มีเส้นผ่านศูนย์กลาง 28-55 นาโนเมตรและเป็นที่ตั้งของเม็ดสีที่ละลายน้ำได้ - ไฟโคบิลิโปรตีนซึ่งกำหนดสีของไซยาโนแบคทีเรียและเกี่ยวข้องกับการสังเคราะห์ด้วยแสง
คลอโรโซมหรือถุงคลอโรเบียมเป็นโครงสร้างที่อุปกรณ์สังเคราะห์แสงของแบคทีเรียสีเขียวในสกุลคลอโรเบียมถูกแปลเป็นภาษาท้องถิ่น มีรูปร่างยาว ยาว 100-150 นาโนเมตร กว้าง 50-70 นาโนเมตร ล้อมรอบด้วยเยื่อหุ้มโปรตีนชั้นเดียว คลอโรโซมอยู่ในชั้นหนาแน่นใต้เยื่อหุ้มเซลล์ไซโตพลาสซึม แต่ถูกแยกออกจากชั้นทางกายภาพ คลอโรโซมของแบคทีเรียสีเขียวประกอบด้วยเม็ดสีสังเคราะห์แสง - แบคทีเรียคลอโรฟิลล์ซึ่งดูดซับควอนตัมแสงและถ่ายโอนพลังงานไปยังศูนย์ปฏิกิริยาของการสังเคราะห์ด้วยแสง
คาร์บอกซีโซม เซลล์ของโปรคาริโอตของโฟโตโทรฟิคบางชนิด (ไซยาโนแบคทีเรีย แบคทีเรียสีม่วงบางชนิด) และโปรคาริโอตที่เป็นเคมีบำบัด (แบคทีเรียไนตริไฟติ้ง) มีโครงสร้างรูปทรงหลายเหลี่ยมขนาด 90-500 นาโนเมตร ตามหน้าที่ที่พวกมันทำ พวกมันเรียกว่าคาร์บอกซีโซม ประกอบด้วยเอนไซม์ไรบูโลส ไดฟอสเฟต คาร์บอกซีเลส ซึ่งกระตุ้นปฏิกิริยาของคาร์บอนไดออกไซด์กับไรบูโลส ไดฟอสเฟต ในวัฏจักรคาลวิน ในแบคทีเรีย autotrophic พวกมันเป็นที่ตั้งของการตรึงคาร์บอนไดออกไซด์ คาร์บอกซีโซมถูกล้อมรอบด้วยเยื่อหุ้มโปรตีนชั้นเดียวซึ่งช่วยปกป้องเอนไซม์จากผลกระทบของโปรตีเอสในเซลล์
สารอาหารสำรอง* นอกเหนือจากองค์ประกอบโครงสร้างที่อธิบายไว้แล้ว ไซโตพลาสซึมของแบคทีเรียยังมีเม็ดที่มีรูปร่างและขนาดต่างๆ ในรูปแบบของสิ่งเจือปน การปรากฏตัวของพวกเขาใน
เซลล์ไม่คงที่และสัมพันธ์กับองค์ประกอบของสารอาหารและสถานะทางสรีรวิทยาของการเพาะเลี้ยง การรวมไซโตพลาสซึมหลายชนิดประกอบด้วยสารประกอบที่ทำหน้าที่เป็นแหล่งพลังงานและแหล่งของสารอาหาร มักก่อตัวขึ้นในการเพาะเลี้ยงบนอาหารสดที่อุดมด้วยสารอาหาร เมื่อการเจริญเติบโตของเซลล์ถูกยับยั้งด้วยเหตุผลบางประการ หรือหลังจากสิ้นสุดระยะเวลาของการเจริญเติบโต องค์ประกอบทางเคมีของสารเจือปนจะแตกต่างกันและไม่เหมือนกันในแบคทีเรียประเภทต่างๆ อาจเป็นโพลีแซ็กคาไรด์ ลิพิด ผลึก และเม็ดของสารอนินทรีย์
ในบรรดาโพลีแซ็กคาไรด์ อันดับแรกเราควรพูดถึงแป้ง ไกลโคเจน และสารคล้ายแป้ง - กรานูโลซา ที่พบมากที่สุดคือไกลโคเจน พบได้ในแบคทีเรีย ซัลโมเนลลา Escherichia coli ปลาซาร์ดีน ฯลฯ ในเซลล์ที่ไม่ใช้ออกซิเจนซึ่งมีสปอร์ในสกุล Clostridium เซลล์จะมีเม็ดแกรนูโลซาขนาดเล็ก เซลล์ใช้สารรวมเหล่านี้เป็นแหล่งพลังงานและคาร์บอน
ไขมันสะสมในไซโตพลาสซึมของแบคทีเรียในรูปของหยดและเมล็ดขนาดเล็ก ในแบคทีเรียหลายชนิด การรวมไขมันจะแสดงด้วยกรดโพลี-พี-ไฮดรอกซีบิวทีริก ซึ่งมักคิดเป็นสัดส่วนมากถึง 50% ของมวลชีวมวลแห้งของแบคทีเรีย แบคทีเรียในสกุล Bacillus และแบคทีเรีย phototrophic อุดมไปด้วยสารประกอบนี้เป็นพิเศษ กรดโพลี-พี-ไฮดรอกซีบิวทีริกถูกสังเคราะห์ในปริมาณมากในระหว่างการเจริญเติบโตของจุลินทรีย์บนตัวกลางที่อุดมไปด้วยคาร์โบไฮเดรต ในแต่ละสายพอลิแลคไทด์ กรด p-ไฮดรอกซีบิวทีริกตกค้างมีมากถึง 60% ดังนั้นสารประกอบนี้จึงเป็น "คลัง" พลังงานในอุดมคติสำหรับแบคทีเรีย จุลินทรีย์บางชนิดสะสมไขและไขมันเป็นกลาง (ไตรกลีเซอไรด์) ดังนั้นในเชื้อมัยโคแบคทีเรียและแอคติโนไมซีตบางครั้งไขจึงมีมากถึง 40% ของมวลแห้ง เซลล์ยีสต์ในสกุล Candida และ Rhodotorula นั้นอุดมไปด้วยไขมันที่เป็นกลางถึงเกือบ 60%
การรวมไขมันทั้งหมดในจุลินทรีย์ทำหน้าที่เป็นแหล่งพลังงานและคาร์บอน
ในเซลล์ของแบคทีเรียหลายชนิด มักพบสารพิเศษที่เรียกว่าเมล็ดโวลูติน โดยธรรมชาติทางเคมี โวลูตินคือโพลีฟอสเฟต ชื่อ โวลูติน
มาจากชื่อสายพันธุ์ของแบคทีเรียกำมะถัน Spirillum volutans ซึ่งได้มีการอธิบายการรวมเหล่านี้ไว้เป็นครั้งแรก Volutin มีคุณสมบัติของ metachromasia เช่น ทำให้สีย้อมบางชนิดเปลี่ยนไป หากแบคทีเรียถูกย้อมด้วยเมทิลีนบลูหรือโทลูอิดีนบลู เม็ดโวลูตินจะกลายเป็นสีม่วงหรือม่วงแดง ในเรื่องนี้ นักวิจัย V. Babes และ E. Ernst ซึ่งเป็นคนแรกที่บรรยายถึงการเจือปนเหล่านี้ เรียกพวกมันว่าเมตาโครมาติกเกรน เม็ด Volutin มีรูปร่างเป็นทรงกลม โดยมีขนาดไม่เกิน 0.5 ไมครอน พวกมันถูกสร้างขึ้นภายใต้สภาวะโภชนาการที่ดีของจุลินทรีย์ โดยเฉพาะอย่างยิ่งในอาหารที่อุดมด้วยคาร์โบไฮเดรต รวมถึงเมื่อมีกลีเซอรอลอยู่ในสิ่งแวดล้อม Volutin พบได้ในเซลล์ของแบคทีเรียที่ทำให้เกิดโรคและ saprophytic เช่น spirilla, azotobacter และสาเหตุของโรคคอตีบ
เซลล์ใช้โวลูตินเป็นแหล่งของกลุ่มฟอสเฟตและพลังงานบางส่วน
ในแบคทีเรียซัลเฟอร์ที่ไม่มีสีและสีม่วงในระหว่างการออกซิเดชั่นของซัลไฟด์แร่ซัลเฟอร์จะสะสมอยู่ในเซลล์ในรูปของหยด การสะสมซัลเฟอร์เกิดขึ้นในสภาพแวดล้อมที่อุดมไปด้วยไฮโดรเจนซัลไฟด์ H2S เมื่อซัลไฟด์หมดไปจากสิ่งแวดล้อม แบคทีเรียจะใช้ซัลเฟอร์ในเซลล์ สำหรับแบคทีเรียซัลเฟอร์ที่ไม่มีสีจะทำหน้าที่เป็นแหล่งพลังงาน สำหรับแบคทีเรียซัลเฟอร์สีม่วงสังเคราะห์ด้วยแสงจะทำหน้าที่เป็นผู้บริจาคอิเล็กตรอน
ในไซยาโนแบคทีเรีย สารสำรองคือไซยาโนไฟซิน เป็นโพลีเปปไทด์ที่ประกอบด้วยอาร์จินีนและกรดแอสปาร์ติก มันทำหน้าที่เป็นแหล่งไนโตรเจนเมื่อขาดในสิ่งแวดล้อม การสะสมของเม็ดไซยาโนไฟซินเกิดขึ้นในระยะการเจริญเติบโตของการเพาะเลี้ยงแบบคงที่และอาจมีน้ำหนักมากถึง 8% ของน้ำหนักแห้งของเซลล์
เยื่อหุ้มเซลล์ไซโตพลาสซึมด้านนอกเป็นฟิล์มบาง ความหนาประมาณ 7-10 นาโนเมตร ภาพยนตร์เรื่องนี้ดูผ่านกล้องจุลทรรศน์อิเล็กตรอนเท่านั้น
โครงสร้าง
องค์ประกอบของไซโตพลาสซึมเมมเบรนมีอะไรบ้าง? โครงสร้างของหนังค่อนข้างหลากหลาย ตามการจัดองค์กรทางเคมี มันเป็นโปรตีนและไขมันที่ซับซ้อน เยื่อหุ้มเซลล์ไซโตพลาสซึมประกอบด้วยชั้นสองชั้น มันทำหน้าที่เป็นฐาน นอกจากนี้เยื่อหุ้มเซลล์ไซโตพลาสซึมยังมีโคเลสเตอรอลและไกลโคลิปิด สารเหล่านี้มีลักษณะเฉพาะโดยแอมพิพาทริซิตี้ กล่าวอีกนัยหนึ่ง พวกมันประกอบด้วยปลายที่ไม่ชอบน้ำ (“กลัวน้ำ”) และปลายที่ชอบน้ำ (“ชอบน้ำ”) ส่วนหลัง (กลุ่มฟอสเฟต) จะถูกส่งออกจากเมมเบรนส่วนหลัง (สารตกค้างจากกรดไขมัน) จะมุ่งเข้าหากัน ด้วยเหตุนี้จึงเกิดชั้นไขมันสองขั้วขึ้น โมเลกุลของไขมันมีความคล่องตัว พวกเขาสามารถเคลื่อนที่ในชั้นเดียวของตัวเองหรือ (ไม่ค่อย) จากที่หนึ่งไปยังอีกที่หนึ่ง
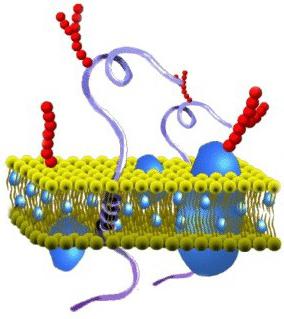
ชั้นไขมันอาจอยู่ในสถานะผลึกแข็งหรือของเหลว Monolayers นั้นไม่สมมาตร ซึ่งหมายความว่าองค์ประกอบของไขมันแตกต่างกัน เนื่องจากคุณสมบัตินี้ เยื่อไซโตพลาสซึมจึงมีความจำเพาะแม้อยู่ภายในเซลล์เดียว องค์ประกอบสำคัญประการที่สองของภาพยนตร์ ได้แก่ โปรตีน การเชื่อมต่อจำนวนมากเหล่านี้สามารถเคลื่อนที่ในระนาบเมมเบรนหรือหมุนรอบแกนของตัวเองได้ อย่างไรก็ตามพวกเขาไม่สามารถย้ายจากส่วนหนึ่งของ bilayer ไปยังอีกส่วนหนึ่งได้ การปกป้องสภาพแวดล้อมภายในเป็นงานหลักที่ดำเนินการโดยเมมเบรนไซโตพลาสซึม นอกจากนี้โครงสร้างของภาพยนตร์ยังช่วยให้มั่นใจได้ถึงกระบวนการต่างๆ โปรตีนมีหน้าที่รับผิดชอบในการทำงานบางอย่าง ลิพิดให้คุณสมบัติเชิงโครงสร้างของฟิล์ม
เยื่อหุ้มเซลล์ไซโตพลาสซึม: หน้าที่
งานหลักคือ:
- สิ่งกีดขวาง- ฟิล์มป้องกันช่วยให้เกิดการแลกเปลี่ยนสารประกอบกับสภาพแวดล้อมภายนอกแบบแอคทีฟ พาสซีฟ เลือกสรร และมีการควบคุม เนื่องจากการซึมผ่านแบบเลือกได้ เซลล์และช่องต่างๆ จะถูกแยกออกจากกันและจัดหาสารที่จำเป็น
- ขนส่ง- การเปลี่ยนผ่านของสารประกอบจากเซลล์หนึ่งไปอีกเซลล์หนึ่งเกิดขึ้นผ่านฟิล์ม ด้วยเหตุนี้จึงมีการส่งมอบสารอาหาร ผลิตภัณฑ์สุดท้ายของการเผาผลาญจะถูกกำจัดออก และสารต่างๆ จะถูกหลั่งออกมา นอกจากนี้ การไล่ระดับสีไอออนิกยังเกิดขึ้น และความเข้มข้นของไอออนิกและ pH จะถูกรักษาไว้ที่ระดับที่เหมาะสมที่สุด จำเป็นต่อการทำงานของเอนไซม์ในเซลล์
งานเสริม

คุณสมบัติพิเศษ
หน้าที่เฉพาะของเมมเบรน ได้แก่ :

ข้อมูลเพิ่มเติม
หากอนุภาคบางตัวไม่สามารถผ่านฟอสโฟไลปิดไบเลเยอร์ได้ไม่ว่าด้วยเหตุผลใดก็ตาม (ตัวอย่างเช่นเนื่องจากคุณสมบัติที่ชอบน้ำเนื่องจากเยื่อหุ้มเซลล์ไซโตพลาสซึมที่อยู่ภายในนั้นไม่ชอบน้ำและไม่อนุญาตให้สารประกอบดังกล่าวผ่านหรือเนื่องจากขนาดใหญ่ ขนาดของอนุภาคเอง) แต่จำเป็น พวกมันสามารถผ่านไปได้ด้วยความช่วยเหลือของโปรตีนตัวพาพิเศษ (ตัวขนส่ง) และโปรตีนแชนเนล หรือการเจาะทะลุผ่านเอ็นโดโทซิส

ในกระบวนการขนส่งแบบพาสซีฟ สารจะข้ามชั้นไขมันโดยการแพร่กระจาย ในกรณีนี้จะไม่สิ้นเปลืองพลังงาน การแพร่กระจายแบบอำนวยความสะดวกอาจเป็นหนึ่งในตัวแปรของกลไกดังกล่าว ในระหว่างกระบวนการนี้ โมเลกุลจำเพาะจะช่วยให้สารผ่านได้สะดวก อาจมีช่องทางที่สามารถผ่านได้เฉพาะอนุภาคชนิดเดียวกันเท่านั้น การขนส่งที่ใช้งานต้องใช้พลังงาน นี่เป็นเพราะความจริงที่ว่ากระบวนการนี้ดำเนินการกับการไล่ระดับความเข้มข้น เมมเบรนไซโตพลาสซึมประกอบด้วยโปรตีนปั๊มพิเศษ รวมถึง ATPase ซึ่งส่งเสริมการเข้าใช้งานของโพแทสเซียมไอออนและการขับถ่ายของไอออนโซเดียม
โมเดล
มีหลายอย่าง:
- "โมเดลแซนวิช"- แนวคิดเรื่องโครงสร้างสามชั้นของเยื่อหุ้มทั้งหมดแสดงโดยนักวิทยาศาสตร์ดอว์สันและดาเนียลีในปี 2478 ในความเห็นของพวกเขา โครงสร้างของภาพยนตร์มีดังนี้ โปรตีน-ไขมัน-โปรตีน ความคิดนี้มีมานานแล้ว
- "โครงสร้างโมเสกเหลว"โมเดลนี้อธิบายโดย Nicholson และ Singer ในปี 1972 ตามนั้นโมเลกุลโปรตีนไม่ได้ก่อตัวเป็นชั้นต่อเนื่อง แต่ถูกแช่อยู่ในชั้นไขมันสองขั้วในรูปแบบของโมเสกจนถึงระดับความลึกที่แตกต่างกัน รุ่นนี้ถือเป็นสากลที่สุด
- "โครงสร้างผลึกโปรตีน".ตามแบบจำลองนี้ เยื่อหุ้มเซลล์ถูกสร้างขึ้นเนื่องจากการรวมตัวกันของโมเลกุลโปรตีนและไขมัน ซึ่งรวมกันบนพื้นฐานของพันธะที่ชอบน้ำและไม่ชอบน้ำ
เยื่อหุ้มเซลล์เรียกอีกอย่างว่าพลาสมา (หรือไซโตพลาสซึม) เมมเบรนและพลาสมาเล็มมา โครงสร้างนี้ไม่เพียงแต่แยกเนื้อหาภายในของเซลล์ออกจากสภาพแวดล้อมภายนอกเท่านั้น แต่ยังเป็นส่วนหนึ่งของออร์แกเนลล์ของเซลล์และนิวเคลียสส่วนใหญ่ด้วย ซึ่งจะแยกพวกมันออกจากไฮยาพลาสซึม (ไซโตซอล) ซึ่งเป็นส่วนของเหลวหนืดของไซโตพลาสซึม ตกลงจะโทรไป. เมมเบรนไซโตพลาสซึมสิ่งที่แยกเนื้อหาของเซลล์ออกจากสภาพแวดล้อมภายนอก เงื่อนไขที่เหลือหมายถึงเมมเบรนทั้งหมด
โครงสร้างของเยื่อหุ้มเซลล์ (ชีวภาพ) ขึ้นอยู่กับไขมัน (ไขมัน) สองชั้น การก่อตัวของชั้นดังกล่าวสัมพันธ์กับลักษณะของโมเลกุล ไขมันไม่ละลายในน้ำ แต่จะควบแน่นด้วยวิธีของมันเอง ส่วนหนึ่งของโมเลกุลไขมันเดี่ยวคือหัวที่มีขั้ว (ถูกดึงดูดโดยน้ำ เช่น ชอบน้ำ) และอีกส่วนหนึ่งเป็นหางยาวที่ไม่มีขั้วคู่หนึ่ง (ส่วนหนึ่งของโมเลกุลนี้ถูกผลักด้วยน้ำ กล่าวคือ ไม่ชอบน้ำ) โครงสร้างของโมเลกุลนี้ทำให้พวกเขา "ซ่อน" หางจากน้ำและหันหัวขั้วโลกไปทางน้ำ
ผลที่ได้คือชั้นไขมันสองชั้น โดยหางที่ไม่มีขั้วอยู่ด้านใน (หันหน้าเข้าหากัน) และส่วนหัวของขั้วอยู่ด้านนอก (หันไปทางสภาพแวดล้อมภายนอกและไซโตพลาสซึม) พื้นผิวของเมมเบรนดังกล่าวเป็นแบบที่ชอบน้ำ แต่ภายในเป็นแบบไม่ชอบน้ำ
ในเยื่อหุ้มเซลล์ ฟอสโฟลิพิดมีชัยเหนือในบรรดาลิพิด (พวกมันอยู่ในลิพิดเชิงซ้อน) หัวของพวกเขามีกรดฟอสฟอริกตกค้าง นอกจากฟอสโฟลิพิดแล้ว ยังมีไกลโคลิพิด (ลิพิด + คาร์โบไฮเดรต) และโคเลสเตอรอล (เกี่ยวข้องกับสเตอรอล) ส่วนหลังให้ความแข็งแกร่งแก่เมมเบรนโดยอยู่ที่ความหนาระหว่างหางของไขมันที่เหลือ (โคเลสเตอรอลไม่เข้ากับน้ำอย่างสมบูรณ์)
เนื่องจากปฏิกิริยาระหว่างไฟฟ้าสถิต โมเลกุลโปรตีนบางชนิดจึงเกาะติดกับหัวไขมันที่มีประจุ ซึ่งกลายเป็นโปรตีนจากเยื่อหุ้มผิว โปรตีนชนิดอื่นๆ ทำปฏิกิริยากับหางที่ไม่มีขั้ว ถูกฝังบางส่วนในชั้นสองชั้น หรือทะลุผ่านหางนั้น
ดังนั้นเยื่อหุ้มเซลล์จึงประกอบด้วยชั้นไขมันสองชั้น พื้นผิว (อุปกรณ์ต่อพ่วง) โปรตีนที่ฝังตัว (กึ่งอินทิกรัล) และโปรตีนที่ซึมผ่าน (อินทิกรัล) นอกจากนี้โปรตีนและไขมันบางชนิดที่อยู่ด้านนอกของเมมเบรนยังสัมพันธ์กับสายโซ่คาร์โบไฮเดรตอีกด้วย

นี้ แบบจำลองโมเสกของเหลวของโครงสร้างเมมเบรนถูกหยิบยกขึ้นมาในยุค 70 ของศตวรรษที่ XX ก่อนหน้านี้ มีการใช้แบบจำลองโครงสร้างแบบแซนด์วิช โดยที่ชั้นไขมัน bilayer ตั้งอยู่ด้านใน และด้านในและด้านนอกของเมมเบรนถูกปกคลุมด้วยชั้นโปรตีนพื้นผิวที่ต่อเนื่องกัน อย่างไรก็ตาม การสะสมข้อมูลการทดลองได้หักล้างสมมติฐานนี้
ความหนาของเยื่อหุ้มเซลล์ต่าง ๆ อยู่ที่ประมาณ 8 นาโนเมตร เมมเบรน (แม้แต่ด้านที่ต่างกันของด้านใดด้านหนึ่ง) จะแตกต่างกันตามเปอร์เซ็นต์ของไขมัน โปรตีน กิจกรรมของเอนไซม์ ฯลฯ ที่แตกต่างกัน เมมเบรนบางชนิดมีของเหลวมากกว่าและซึมผ่านได้มากกว่า เมมเบรนบางชนิดมีความหนาแน่นมากกว่า
เยื่อหุ้มเซลล์แตกตัวรวมกันได้ง่ายเนื่องจากคุณสมบัติทางเคมีกายภาพของชั้นไขมัน ในระนาบของเมมเบรน ไขมันและโปรตีน (เว้นแต่ว่าพวกมันจะถูกยึดโดยโครงร่างโครงกระดูก) จะเคลื่อนไหว
หน้าที่ของเยื่อหุ้มเซลล์
โปรตีนส่วนใหญ่ที่แช่อยู่ในเยื่อหุ้มเซลล์ทำหน้าที่ของเอนไซม์ (คือเอนไซม์)
บ่อยครั้ง (โดยเฉพาะในเยื่อหุ้มเซลล์ของออร์แกเนลล์) เอนไซม์อยู่ในลำดับที่แน่นอนเพื่อให้ผลิตภัณฑ์ปฏิกิริยาที่ถูกเร่งปฏิกิริยาโดยเอนไซม์ตัวหนึ่งเคลื่อนที่ไปยังตัวที่สองจากนั้นตัวที่สามเป็นต้น เครื่องลำเลียงถูกสร้างขึ้นเพื่อรักษาเสถียรภาพของโปรตีนบนพื้นผิวเนื่องจากไม่ได้ ปล่อยให้เอนไซม์ลอยไปตามชั้นไลปิด
เยื่อหุ้มเซลล์ทำหน้าที่กำหนดขอบเขต (สิ่งกีดขวาง) จากสิ่งแวดล้อมและในขณะเดียวกันก็ทำหน้าที่ขนส่ง
เราสามารถพูดได้ว่านี่คือจุดประสงค์ที่สำคัญที่สุด เมมเบรนไซโตพลาสซึมซึ่งมีความแข็งแรงและความสามารถในการซึมผ่านแบบเลือกได้ช่วยรักษาความคงที่ขององค์ประกอบภายในของเซลล์ (สภาวะสมดุลและความสมบูรณ์ของมัน)
ในกรณีนี้การขนส่งสารเกิดขึ้นได้หลายวิธี การขนส่งตามระดับความเข้มข้นเกี่ยวข้องกับการเคลื่อนที่ของสารจากบริเวณที่มีความเข้มข้นสูงกว่าไปยังบริเวณที่มีความเข้มข้นต่ำกว่า (การแพร่กระจาย) ตัวอย่างเช่น ก๊าซ (CO 2 , O 2 ) แพร่กระจาย
นอกจากนี้ยังมีการขนส่งแบบไล่ระดับความเข้มข้นด้วย แต่มีการใช้พลังงาน
โปรตีนที่ร้อยเกลียวรวมกันเพื่อสร้างรูพรุนสำหรับการเคลื่อนที่ของสารบางชนิดผ่านเมมเบรน พาหะดังกล่าวไม่เคลื่อนที่ แต่ก่อตัวเป็นช่องทางในเมมเบรนและทำงานคล้ายกับเอนไซม์ซึ่งจับกับสารเฉพาะ การถ่ายโอนเกิดขึ้นเนื่องจากการเปลี่ยนแปลงโครงสร้างโปรตีน ส่งผลให้เกิดการสร้างช่องในเมมเบรน ตัวอย่างคือปั๊มโซเดียมโพแทสเซียม
ฟังก์ชั่นการขนส่งของเยื่อหุ้มเซลล์ยูคาริโอตนั้นเกิดขึ้นได้ผ่านทางเอนโดไซโตซิส (และเอ็กโซไซโทซิส)ด้วยกลไกเหล่านี้ โมเลกุลขนาดใหญ่ของพอลิเมอร์ชีวภาพ แม้กระทั่งเซลล์ทั้งหมด จึงสามารถเข้าสู่เซลล์ (และออกจากเซลล์ได้) Endo และ exocytosis ไม่ใช่ลักษณะของเซลล์ยูคาริโอตทั้งหมด (โปรคาริโอตไม่มีเลย) ดังนั้นการสังเกต endocytosis ในโปรโตซัวและสัตว์ไม่มีกระดูกสันหลังส่วนล่าง ในสัตว์เลี้ยงลูกด้วยนมเม็ดเลือดขาวและมาโครฟาจจะดูดซับสารและแบคทีเรียที่เป็นอันตรายเช่น endocytosis ทำหน้าที่ป้องกันร่างกาย
Endocytosis แบ่งออกเป็น ฟาโกไซโตซิส(ไซโตพลาสซึมห่อหุ้มอนุภาคขนาดใหญ่) และ พิโนไซโทซิส(จับหยดของเหลวที่มีสารที่ละลายอยู่ในนั้น) กลไกของกระบวนการเหล่านี้ใกล้เคียงกัน สารที่ถูกดูดซับบนพื้นผิวของเซลล์จะถูกล้อมรอบด้วยเมมเบรน เกิดตุ่ม (phagocytic หรือ pinocytic) ซึ่งจะเคลื่อนเข้าสู่เซลล์
Exocytosis คือการกำจัดสารออกจากเซลล์ (ฮอร์โมน โพลีแซ็กคาไรด์ โปรตีน ไขมัน ฯลฯ) โดยเยื่อหุ้มเซลล์ สารเหล่านี้มีอยู่ในถุงเมมเบรนที่พอดีกับเยื่อหุ้มเซลล์ เยื่อหุ้มทั้งสองผสานกันและเนื้อหาปรากฏอยู่นอกเซลล์
เมมเบรนไซโตพลาสซึมทำหน้าที่รับเมื่อต้องการทำเช่นนี้ โครงสร้างจะอยู่ที่ด้านนอกซึ่งสามารถรับรู้ถึงสิ่งกระตุ้นทางเคมีหรือกายภาพ โปรตีนบางชนิดที่ทะลุพลาสมาเลมมานั้นเชื่อมต่อจากด้านนอกเข้ากับโซ่โพลีแซ็กคาไรด์ (ก่อตัวเป็นไกลโคโปรตีน) เหล่านี้คือตัวรับโมเลกุลที่แปลกประหลาดซึ่งจับฮอร์โมน เมื่อฮอร์โมนตัวใดตัวหนึ่งจับกับตัวรับ มันจะเปลี่ยนโครงสร้างของมัน ซึ่งจะกระตุ้นให้เกิดกลไกการตอบสนองของเซลล์ ในกรณีนี้ ช่องสามารถเปิดได้ และสารบางชนิดสามารถเริ่มเข้าหรือออกจากเซลล์ได้
การทำงานของตัวรับของเยื่อหุ้มเซลล์ได้รับการศึกษาอย่างดีโดยพิจารณาจากการกระทำของฮอร์โมนอินซูลิน เมื่ออินซูลินจับกับตัวรับไกลโคโปรตีน ตัวเร่งปฏิกิริยาภายในเซลล์ของโปรตีนนี้ (เอนไซม์อะดีนิเลตไซคลอส) จะถูกกระตุ้น เอนไซม์สังเคราะห์ cyclic AMP จาก ATP มันกระตุ้นหรือยับยั้งเอนไซม์ต่างๆ ของการเผาผลาญของเซลล์แล้ว
การทำงานของตัวรับของเมมเบรนไซโตพลาสซึมยังรวมถึงการจดจำเซลล์ข้างเคียงที่เป็นชนิดเดียวกันด้วย เซลล์ดังกล่าวเกาะติดกันด้วยการสัมผัสระหว่างเซลล์ต่างๆ
ในเนื้อเยื่อด้วยความช่วยเหลือของการสัมผัสระหว่างเซลล์ เซลล์สามารถแลกเปลี่ยนข้อมูลระหว่างกันโดยใช้สารโมเลกุลต่ำที่สังเคราะห์ขึ้นเป็นพิเศษ ตัวอย่างหนึ่งของปฏิสัมพันธ์ดังกล่าวคือการยับยั้งการสัมผัส เมื่อเซลล์หยุดการเจริญเติบโตหลังจากได้รับข้อมูลว่าพื้นที่ว่างถูกครอบครอง
การติดต่อระหว่างเซลล์สามารถทำได้ง่าย (เยื่อหุ้มเซลล์ต่าง ๆ ติดกัน), การล็อค (การบุกรุกของเยื่อหุ้มเซลล์หนึ่งไปยังอีกเซลล์หนึ่ง), เดสโมโซม (เมื่อเยื่อหุ้มเชื่อมต่อกันด้วยมัดของเส้นใยตามขวางที่เจาะเข้าไปในไซโตพลาสซึม) นอกจากนี้ยังมีการติดต่อระหว่างเซลล์หลายแบบเนื่องจากผู้ไกล่เกลี่ย (ตัวกลาง) - ไซแนปส์ ในนั้นสัญญาณจะถูกส่งไม่เพียงแต่ทางเคมีเท่านั้น แต่ยังส่งสัญญาณทางไฟฟ้าด้วย ไซแนปส์ส่งสัญญาณระหว่างเซลล์ประสาท เช่นเดียวกับจากเส้นประสาทไปยังเซลล์กล้ามเนื้อ
เยื่อหุ้มเซลล์ไซโตพลาสซึมประกอบด้วยสามชั้น:
ภายนอก – โปรตีน;
ไขมันชั้นกลาง - สองโมเลกุล;
ภายใน - โปรตีน
ความหนาของเมมเบรนคือ 7.5-10 นาโนเมตร ชั้นไขมันสองโมเลกุลเป็นเมทริกซ์ของเมมเบรน โมเลกุลไขมันของทั้งสองชั้นมีปฏิกิริยากับโมเลกุลโปรตีนที่แช่อยู่ในนั้น ไขมันเมมเบรนจาก 60 ถึง 75% เป็นฟอสโฟลิปิด, 15-30% เป็นคอเลสเตอรอล โปรตีนส่วนใหญ่จะแสดงโดยไกลโคโปรตีน แยกแยะ โปรตีนอินทิกรัลซึมซับเมมเบรนทั้งหมดและ อุปกรณ์ต่อพ่วงตั้งอยู่บนพื้นผิวด้านนอกหรือด้านใน
โปรตีนอินทิกรัลสร้างช่องไอออนเพื่อให้แน่ใจว่ามีการแลกเปลี่ยนไอออนบางอย่างระหว่างของเหลวภายนอกและภายในเซลล์ พวกมันยังเป็นเอ็นไซม์ที่ทำหน้าที่ขนส่งไอออนผ่านเมมเบรนแบบทวนระดับ
โปรตีนส่วนปลายเป็นตัวรับสารเคมีบนพื้นผิวด้านนอกของเมมเบรนที่สามารถโต้ตอบกับสารออกฤทธิ์ทางสรีรวิทยาต่างๆ
ฟังก์ชั่นเมมเบรน:
1. รับประกันความสมบูรณ์ของเซลล์ในฐานะหน่วยโครงสร้างของเนื้อเยื่อ
ดำเนินการแลกเปลี่ยนไอออนระหว่างไซโตพลาสซึมและของเหลวนอกเซลล์
ช่วยลำเลียงไอออนและสารอื่นๆ เข้าและออกจากเซลล์
ดำเนินการรับรู้และประมวลผลข้อมูลที่มาถึงเซลล์ในรูปแบบของสัญญาณทางเคมีและไฟฟ้า
กลไกการกระตุ้นเซลล์ ประวัติความเป็นมาของการศึกษาปรากฏการณ์ไฟฟ้าชีวภาพ
ข้อมูลส่วนใหญ่ที่ส่งผ่านในร่างกายจะอยู่ในรูปแบบของสัญญาณไฟฟ้า (เช่น แรงกระตุ้นของเส้นประสาท) การมีอยู่ของกระแสไฟฟ้าจากสัตว์ก่อตั้งขึ้นครั้งแรกโดยนักวิทยาศาสตร์ธรรมชาติ (นักสรีรวิทยา) แอล. กัลวานี ในปี พ.ศ. 2329 เพื่อศึกษาไฟฟ้าในชั้นบรรยากาศ เขาได้ระงับการเตรียมขากบไว้บนตะขอทองแดง เมื่ออุ้งเท้าสัมผัสกับราวเหล็กของระเบียง กล้ามเนื้อก็เกิดการหดตัว สิ่งนี้บ่งบอกถึงการกระทำของกระแสไฟฟ้าบางชนิดบนเส้นประสาทของยาประสาทและกล้ามเนื้อ กัลวานีเชื่อว่าสิ่งนี้เกิดจากการมีไฟฟ้าอยู่ในเนื้อเยื่อของสิ่งมีชีวิตนั่นเอง อย่างไรก็ตาม A. Volta กำหนดว่าแหล่งกำเนิดไฟฟ้าเป็นสถานที่สัมผัสของโลหะสองชนิดที่แตกต่างกัน - ทองแดงและเหล็ก ในด้านสรีรวิทยา การทดลองคลาสสิกครั้งแรกของกัลวานีถือเป็นการสัมผัสเส้นประสาทของการเตรียมประสาทและกล้ามเนื้อด้วยแหนบ bimetallic ที่ทำจากทองแดงและเหล็ก เพื่อพิสูจน์ว่าเขาพูดถูก กัลวานีจึงจัดทำ ประสบการณ์ครั้งที่สอง- เขาโยนปลายเส้นประสาทที่กระตุ้นการเตรียมประสาทและกล้ามเนื้อไปบนรอยตัดของกล้ามเนื้อ ส่งผลให้มันลดลง อย่างไรก็ตาม ประสบการณ์นี้ไม่ได้โน้มน้าวคนรุ่นเดียวกันของกัลวานี ดังนั้น Matteuci ชาวอิตาลีอีกคนจึงทำการทดลองต่อไปนี้ เขาวางเส้นประสาทของกบตัวหนึ่งที่เตรียมประสาทและกล้ามเนื้อไว้บนกล้ามเนื้อของกบตัวที่สอง ซึ่งหดตัวภายใต้อิทธิพลของกระแสที่น่ารำคาญ ส่งผลให้ยาตัวแรกเริ่มหดตัวด้วย สิ่งนี้บ่งบอกถึงการถ่ายโอนไฟฟ้า (ศักยภาพในการดำเนินการ) จากกล้ามเนื้อหนึ่งไปยังอีกกล้ามเนื้อหนึ่ง การมีอยู่ของความแตกต่างที่อาจเกิดขึ้นระหว่างบริเวณที่เสียหายและไม่เสียหายของกล้ามเนื้อนั้นเกิดขึ้นอย่างแม่นยำครั้งแรกในศตวรรษที่ 19 โดยใช้เครื่องวัดกระแสไฟฟ้าแบบสตริง (แอมป์มิเตอร์) โดย Matteuci นอกจากนี้ บาดแผลยังมีประจุลบ และพื้นผิวของกล้ามเนื้อมี ประจุบวก