
Aké vlastnosti má cytoplazmatická membrána? Bunková membrána
Vonkajšia cytoplazmatická membrána je tenký film. Jeho hrúbka je asi 7-10 nm. Film je možné pozorovať iba cez elektrónový mikroskop.
Štruktúra
Aké je zloženie cytoplazmatickej membrány? Štruktúra filmu je dosť rôznorodá. Podľa chemickej organizácie ide o komplex bielkovín a lipidov. Cytoplazmatická membrána bunky obsahuje dvojvrstvu. Pôsobí ako základ. Okrem toho cytoplazmatická membrána obsahuje cholesterol a glykolipidy. Tieto látky sa vyznačujú amfipatričnosťou. Inými slovami, obsahujú hydrofóbne („vody sa obávajúce“) a hydrofilné („vodu milujúce“) konce. Posledne menované (fosfátová skupina) sú nasmerované smerom von z membrány, druhé (zvyšky mastných kyselín) sú orientované k sebe. Vďaka tomu sa vytvára lipidová bipolárna vrstva. Molekuly lipidov majú pohyblivosť. Sú schopní pohybovať sa vo vlastnej monovrstve alebo (zriedkavo) z jednej do druhej.

Lipidová vrstva môže byť v pevnom alebo tekutom kryštálovom stave. Monovrstvy sú asymetrické. To znamená, že ich lipidové zloženie je odlišné. Vďaka tejto vlastnosti majú cytoplazmatické membrány špecifickosť aj v rámci jednej bunky. Druhá podstatná zložka filmu zahŕňa proteíny. Mnohé z týchto spojení sa môžu pohybovať v rovine membrány alebo rotovať okolo vlastnej osi. Nie sú však schopné prechádzať z jednej časti dvojvrstvy do druhej. Ochrana vnútorného prostredia je hlavnou úlohou, ktorú vykonáva cytoplazmatická membrána. Štruktúra filmu navyše zabezpečuje priebeh rôznych procesov. Proteíny sú zodpovedné za vykonávanie určitých úloh. Lipidy poskytujú štrukturálne vlastnosti filmu.
Cytoplazmatická membrána: funkcie
Hlavnými úlohami sú:
- Bariéra. Ochranný film zabezpečuje aktívnu, pasívnu, selektívnu, regulovanú výmenu zlúčenín s vonkajším prostredím. Vďaka selektívnej priepustnosti sú bunka a jej oddelenia oddelené a zásobované potrebnými látkami.
- Doprava. Prechod zlúčenín z bunky do bunky prebieha cez film. Vďaka tomu sa dodávajú nutričné zlúčeniny, odstraňujú sa konečné produkty metabolizmu a vylučujú sa rôzne látky. Okrem toho sa vytvárajú iónové gradienty a koncentrácia iónov a pH sa udržiavajú na optimálnych úrovniach. Sú nevyhnutné pre aktívnu činnosť bunkových enzýmov.
Pomocné úlohy

Špeciálne vlastnosti
Medzi špecifické funkcie membrány patria:

Ďalšie informácie
Ak niektoré častice z jedného alebo druhého dôvodu nie sú schopné prejsť cez fosfolipidovú dvojvrstvu (napríklad kvôli hydrofilným vlastnostiam, pretože cytoplazmatická membrána vo vnútri je hydrofóbna a neumožňuje takýmto zlúčeninám prejsť, alebo kvôli veľkej veľkosť samotných častíc), ale sú nevyhnutné, potom môžu prechádzať pomocou špeciálnych nosných proteínov (transportérov) a kanálových proteínov. Alebo sa ich penetrácia uskutočňuje endocytózou.
V procese pasívneho transportu látky prechádzajú cez lipidovú vrstvu difúziou. V tomto prípade nedochádza k plytvaniu energiou. Uľahčená difúzia môže byť jedným z variantov takéhoto mechanizmu. Počas tohto procesu špecifická molekula uľahčuje prechod látky. Môže mať kanál schopný prechádzať len časticami rovnakého typu. Aktívny transport spotrebúva energiu. Je to spôsobené tým, že tento proces sa uskutočňuje proti koncentračnému gradientu. Cytoplazmatická membrána obsahuje špeciálne pumpové proteíny vrátane ATPázy, ktorá podporuje aktívny vstup iónov draslíka a vylučovanie iónov sodíka.
Modelky
Je ich niekoľko:
- "Sendvičový model". Myšlienku trojvrstvovej štruktúry všetkých membrán vyjadrili vedci Dawson a Danieli v roku 1935. Podľa ich názoru bola štruktúra filmu nasledovná: proteíny-lipidy-proteíny. Táto myšlienka existovala pomerne dlho.
- "Štruktúra tekutej mozaiky." Tento model opísali Nicholson a Singer v roku 1972. V súlade s ním molekuly proteínov netvoria súvislú vrstvu, ale sú ponorené do bipolárnej lipidovej vrstvy vo forme mozaiky do rôznej hĺbky. Tento model je považovaný za najuniverzálnejší.
- "Kryštálová štruktúra proteínu". Podľa tohto modelu vznikajú membrány v dôsledku prelínania molekúl proteínov a lipidov, ktoré sa spájajú na základe hydrofilno-hydrofóbnych väzieb.
Každá živá bunka je oddelená od prostredia tenkou membránou špeciálnej štruktúry - cytoplazmatickou membránou (CPM). Eukaryoty majú početné intracelulárne membrány, ktoré oddeľujú priestor organel od cytoplazmy, zatiaľ čo pre väčšinu prokaryotov je CPM jedinou bunkovou membránou. V niektorých baktériách a archeách môže prenikať do cytoplazmy, pričom vytvára výrastky a záhyby rôznych tvarov.
CPM akýchkoľvek buniek sú postavené podľa jediného plánu a pozostávajú z fosfolipidov (obr. 3.5, A). V baktériách obsahujú dve mastné kyseliny, zvyčajne so 16-18 atómami uhlíka v reťazci a s nasýtenou alebo jednou nenasýtenou väzbou, spojené esterovou väzbou s dvoma hydroxylovými skupinami glycerolu. Zloženie mastných kyselín baktérií sa môže meniť v závislosti od zmien prostredia, najmä teploty. Pri znižovaní teploty sa zvyšuje množstvo nenasýtených mastných kyselín v zložení fosfolipidov, čo výrazne ovplyvňuje tekutosť membrány. Niektoré mastné kyseliny môžu byť rozvetvené alebo môžu obsahovať cyklopropánový kruh. Tretia OH skupina glycerolu je pripojená k zvyšku kyseliny fosforečnej a cez ňu k hlavnej skupine. Hlavné skupiny fosfolipidov môžu mať u rôznych prokaryotov rôznu chemickú povahu (fosfatidyletanolamín, fosfatidylglycerol, kardiolipín, fosfatidylserín, lecitín atď.), ale majú jednoduchšiu štruktúru ako u eukaryotov. Napríklad pri E. coli, sú zastúpené 75 % fosfatidyletanolamínom, 20 % fosfatidylglycerolom, zvyšok tvoria kardiolipín (difosfatidylglycerol), fosfatidylserín a stopové množstvá iných zlúčenín. Iné baktérie majú zložitejšie typy membránových lipidov. Niektoré bunky tvoria glykolipidy, ako je monogalaktozyldiglycerid. Lipidy archaálnej membrány sa líšia od eukaryotických a bakteriálnych. Namiesto mastných kyselín obsahujú vyššie izoprenoidové alkoholy naviazané na glycerol skôr jednoduchou ako esterovou väzbou.
Ryža. 3.5.
A- fosfolipid; b- dvojvrstvová membrána
O O o O o o
Takéto molekuly tvoria membránovú dvojvrstvu, kde hydrofóbne časti smerujú dovnútra a hydrofilné časti smerom von do prostredia a do cytoplazmy (obr. 3.5, Obr. b). Mnohé proteíny sú zabudované v dvojvrstve alebo ju pretínajú a môžu difundovať v membráne, pričom niekedy vytvárajú komplexné komplexy. Membránové proteíny majú množstvo dôležitých funkcií, medzi ktoré patrí premena a ukladanie metabolickej energie, regulácia vstrebávania a uvoľňovania všetkých živín a metabolických produktov. Okrem toho rozpoznávajú a prenášajú mnohé signály odrážajúce zmeny v prostredí a spúšťajú zodpovedajúcu kaskádu reakcií vedúcich k bunkovej odpovedi. Táto organizácia membrán je dobre vysvetlená modelom tekutých kryštálov s mozaikou rozptýlenou membránovými proteínmi (obr. 3.6).

Ryža. 3.6.
Väčšina biologických membrán má hrúbku 4 až 7 nm. Bunkové membrány sú jasne viditeľné v transmisnom elektrónovom mikroskope v porovnaní s ťažkými kovmi. Na elektrónových mikrosnímkach vyzerajú ako trojvrstvové útvary: dve vonkajšie tmavé vrstvy ukazujú polohu polárnych skupín lipidov a svetlá stredná vrstva znázorňuje hydrofóbny vnútorný priestor (obr. 3.7).
Ďalšou technikou na štúdium membrán je získanie rozštiepených buniek zmrazených pri teplote tekutého dusíka a kontrast výsledných povrchov naprašovaním ťažkých kovov.
(platina, zlato, striebro). Výsledné prípravky sa prezerajú pod skenovacím elektrónovým mikroskopom. V tomto prípade je možné vidieť povrch membrány a v nej obsiahnuté mozaikové membránové proteíny, ktoré neprechádzajú cez membránu, ale sú spojené špeciálnymi oblasťami hydrofóbnej kotvy s hydrofóbnou oblasťou dvojvrstvy.

Ryža. 3.7.
CPM má vlastnosť selektívnej permeability, ktorá bráni voľnému pohybu väčšiny látok do bunky a von z bunky a tiež hrá významnú úlohu pri raste a delení buniek, pohybe a exporte povrchových a extracelulárnych proteínov a sacharidov (exopolysacharidov). . Ak je bunka umiestnená v prostredí s vyšším alebo nižším osmotickým tlakom ako vo vnútri cytoplazmy, potom voda bunku opustí alebo do nej vnikne. To odráža vlastnosť vody vyrovnávať gradienty roztoku. V tomto prípade sa cytoplazma zmršťuje alebo expanduje (fenomén plazmolýzy/deplazmolýzy). Väčšina baktérií však pri takýchto experimentoch nemení svoj tvar kvôli prítomnosti tuhej bunkovej steny.
CPM reguluje tok živín a metabolitov. Prítomnosť hydrofóbnej vrstvy tvorenej membránovými lipidmi zabraňuje prechodu akýchkoľvek polárnych molekúl a makromolekúl cez ňu. Táto vlastnosť umožňuje bunkám, ktoré vo všeobecnosti existujú v zriedených roztokoch, zachovať si užitočné makromolekuly a metabolické prekurzory. Bunková membrána je tiež navrhnutá tak, aby vykonávala transportnú funkciu. Typicky majú prokaryoty veľký počet veľmi špecifických transportných systémov. Transport je integrálnou súčasťou celkovej bioenergetiky bunky, ktorá prostredníctvom CPM vytvára a využíva rôzne iónové gradienty na transport látok a vytváranie ďalších gradientov potrebných pre bunku. CPM hrá významnú úlohu pri pohybe, raste a delení buniek. Mnohé metabolické procesy sú sústredené v membráne prokaryotov. Membránové proteíny plnia dôležité funkcie: podieľajú sa na premene a skladovaní energie, regulujú vstrebávanie a uvoľňovanie všetkých živín a produktov látkovej premeny, rozpoznávajú a prenášajú signály o zmenách prostredia.
Vonkajšia cytoplazmatická membrána obklopujúca cytoplazmu každej bunky určuje jej veľkosť a zabezpečuje udržiavanie významných rozdielov medzi bunkovým obsahom a prostredím. Membrána slúži ako vysoko selektívny filter, ktorý udržuje rozdiel v koncentráciách iónov na oboch stranách membrány a umožňuje živinám prenikať do bunky a odpadovým látkam opúšťať bunku.
Všetky biologické membrány sú zostavy molekúl lipidov a proteínov, ktoré sú držané pohromade nekovalentnými interakciami. Molekuly lipidov a proteínov tvoria súvislú dvojitú vrstvu.
Lipidová dvojvrstva je hlavnou štruktúrou membrány, ktorá vytvára relatívne nepriepustnú bariéru pre väčšinu molekúl rozpustných vo vode.
Proteínové molekuly sú akoby „rozpustené“ v lipidovej dvojvrstve. Prostredníctvom proteínov sa vykonávajú rôzne funkcie membrány: niektoré z nich zabezpečujú transport určitých molekúl do bunky alebo z bunky, iné sú enzýmy a katalyzujú reakcie spojené s membránou a iné zabezpečujú štrukturálne spojenie medzi cytoskeletom a extracelulárnou matricou. alebo slúžia ako receptory na príjem a konverziu chemických signálov z prostredia.
Dôležitou vlastnosťou biologických membrán je tekutosť. Všetky bunkové membrány sú pohyblivé tekuté štruktúry: väčšina ich základných lipidových a proteínových molekúl je schopná sa pomerne rýchlo pohybovať v rovine membrány. Ďalšou vlastnosťou membrán je ich asymetria: obe ich vrstvy sa líšia lipidovým a proteínovým zložením, čo odráža funkčné rozdiely ich povrchov.
Funkcie vonkajšej cytoplazmatickej membrány:
· bariéra – zabezpečuje regulovaný, selektívny, pasívny a aktívny metabolizmus s okolím. Selektívna permeabilita zabezpečuje, že bunka a bunkové kompartmenty sú oddelené od prostredia a zásobené potrebnými látkami.
Transport - transport látok do bunky az bunky prebieha cez membránu. Transport cez membrány zabezpečuje: dodávanie živín, odstraňovanie konečných produktov metabolizmu, sekréciu rôznych látok, vytváranie iónových gradientov, udržiavanie vhodného pH a koncentrácie iónov v bunke, ktoré sú potrebné pre fungovanie bunkových enzýmov.
Častice, ktoré z nejakého dôvodu nie sú schopné prejsť cez fosfolipidovú dvojvrstvu (napríklad kvôli hydrofilným vlastnostiam, keďže membrána vo vnútri je hydrofóbna a neprepúšťa hydrofilné látky, alebo kvôli ich veľkým rozmerom), ale sú nevyhnutné pre bunka, môže preniknúť cez membránu cez špeciálne nosné proteíny (transportéry) a kanálové proteíny alebo endocytózou.
Počas pasívneho transportu látky prechádzajú cez lipidovú dvojvrstvu bez spotreby energie, difúziou. Variantom tohto mechanizmu je uľahčená difúzia, pri ktorej špecifická molekula pomáha látke prejsť cez membránu. Táto molekula môže mať kanál, ktorý umožňuje prechod len jedného typu látky.
Aktívny transport vyžaduje energiu, pretože sa vyskytuje proti koncentračnému gradientu. Na membráne sú špeciálne pumpové proteíny vrátane ATPázy, ktoré aktívne pumpujú draselné ióny (K+) do bunky a pumpujú z nej sodíkové ióny (Na+).
· matrica – zabezpečuje určitú relatívnu polohu a orientáciu membránových proteínov, ich optimálnu interakciu;
· mechanická – zabezpečuje autonómiu bunky, jej vnútrobunkových štruktúr, ako aj spojenie s inými bunkami (v tkanivách). Bunkové steny zohrávajú hlavnú úlohu pri zabezpečovaní mechanickej funkcie a u zvierat medzibunková látka.
· energia - pri fotosyntéze v chloroplastoch a bunkovom dýchaní v mitochondriách fungujú v ich membránach systémy prenosu energie, na ktorých sa podieľajú aj bielkoviny;
· receptor – niektoré proteíny sediace v membráne sú receptory (molekuly, pomocou ktorých bunka vníma určité signály).
Napríklad hormóny cirkulujúce v krvi pôsobia len na cieľové bunky, ktoré majú receptory zodpovedajúce týmto hormónom. Neurotransmitery (chemické látky, ktoré zabezpečujú vedenie nervových vzruchov) sa tiež viažu na špeciálne receptorové proteíny v cieľových bunkách.
· enzymatické – membránové proteíny sú často enzýmy. Napríklad plazmatické membrány buniek črevného epitelu obsahujú tráviace enzýmy.
· implementácia tvorby a vedenia biopotenciálov.
Pomocou membrány sa v bunke udržiava konštantná koncentrácia iónov: koncentrácia iónu K+ vo vnútri bunky je oveľa vyššia ako vonku a koncentrácia Na+ je oveľa nižšia, čo je veľmi dôležité, pretože to zaisťuje udržiavanie rozdielu potenciálov na membráne a generovanie nervového impulzu.
· značenie buniek – na membráne sú antigény, ktoré fungujú ako markery – „štítky“, ktoré umožňujú bunku identifikovať. Sú to glykoproteíny (t. j. proteíny s rozvetvenými bočnými oligosacharidovými reťazcami, ktoré sú k nim pripojené), ktoré zohrávajú úlohu „antén“. Kvôli nespočetným konfiguráciám bočných reťazcov je možné vytvoriť špecifický marker pre každý typ bunky. Pomocou markerov môžu bunky rozpoznať iné bunky a konať v zhode s nimi, napríklad pri tvorbe orgánov a tkanív. To tiež umožňuje imunitnému systému rozpoznať cudzie antigény.
Každé ľudské alebo zvieracie telo pozostáva z miliárd buniek. Bunka je komplexný mechanizmus, ktorý vykonáva špecifické funkcie. Všetky orgány a tkanivá pozostávajú z podjednotiek.
Systém má cytoplazmatickú membránu, cytoplazmu, jadro a množstvo organel. Jadro je oddelené od organel vnútornou membránou. Všetky spolu poskytujú život tkanivám a tiež umožňujú metabolizmus.
Vo fungovaní hrá dôležitú úlohu cytoplazmatická plazmatická lemma alebo membrána.
Samotný názov, vonkajšia cytoplazmatická membrána, pochádza z latinského membrana, alebo inak koža. Toto je oddeľovač priestoru medzi bunkovými organizmami.
Hypotéza štruktúry bola predložená už v roku 1935. V roku 1959 V. Robertson dospel k záveru, že membránové plášte sú usporiadané podľa rovnakého princípu.
Vďaka veľkému množstvu nahromadených informácií získala dutina tekutý mozaikový model štruktúry. Teraz sa to považuje za všeobecne akceptované. Je to vonkajšia cytoplazmatická membrána, ktorá tvorí vonkajší obal jednotiek.
Štruktúra
Čo je teda plazmová lemma?
Je to tenký film oddeľujúci prokaryoty od vnútorného prostredia. Dá sa to vidieť iba cez mikroskop. Štruktúra cytoplazmatickej membrány zahŕňa dvojvrstvu, ktorá slúži ako základ.
Dvojvrstva je dvojitá vrstva pozostávajúca z proteínov a lipidov. Existujú aj cholesterol a glykolipidy, ktoré sú amfipatričné.
Čo to znamená?
Tukový organizmus má bipolárnu hlavu a hydrofilný chvost. Prvý je spôsobený strachom z vody a druhý je spôsobený jej absorpciou. Skupina fosfátov má smerom von z filmu, pričom tieto sú nasmerované k sebe.
Tak sa vytvorí bipolárna lipidová vrstva. Lipidy sú vysoko aktívne, môžu sa pohybovať vo svojej monovrstve a zriedka sa presúvajú do iných oblastí.
Polyméry sa delia na:
- vonkajší,
- integrálne,
- prestupujúcich plazmovú lemu.
Prvé sú umiestnené iba na povrchovej časti sínusu. Sú držané pohromade elektrostatikou s bipolárnymi hlavami lipidových prvkov. Zachováva nutričné enzýmy. Integrálne vo vnútri, sú zabudované do samotnej štruktúry plášťa, spojenia menia svoju polohu v dôsledku pohybu eukaryotov. Slúžia ako druh dopravníka, postaveného tak, že po nich prúdia substráty a produkty reakcie. Proteínové zlúčeniny prenikajúce do makrodutiny majú vlastnosti tvorby pórov pre vstup živín do tela.
Core
Každá jednotka má jadro, to je jej základ. Cytoplazmatická membrána má tiež organelu, ktorej štruktúra bude opísaná nižšie.
Jadrová štruktúra zahŕňa membránu, miazgu, miesto zostavenia ribozómov a chromatín. Plášť je rozdelený jadrovým priestorom, je obklopený kvapalinou.
Funkcie organely sú rozdelené do dvoch hlavných:
- uzavretie štruktúry v organele,
- regulácia obsahu jadra a kvapaliny.
Jadro pozostáva z pórov, z ktorých každý je určený prítomnosťou kombinácií ťažkých pórov. Ich objem môže naznačovať aktívnu motorickú schopnosť eukaryotov. Napríklad vysokoaktívne nezrelé látky obsahujú väčší počet oblastí pórov. Proteíny slúžia ako jadrová šťava.
Polyméry predstavujú kombináciu matrice a nukleoplazmy. Kvapalina je obsiahnutá vo vnútri jadrového filmu a zabezpečuje funkčnosť genetického obsahu organizmov. Proteínový prvok poskytuje ochranu a silu podjednotkám.
Ribozomálne RNA dozrievajú v samotnom jadierku. Samotné gény RNA sa nachádzajú v špecifickej oblasti niekoľkých chromozómov. V rámci nich sa formujú malí organizátori. Vo vnútri sa vytvárajú samotné jadierka. Zóny v mitotických chromozómoch sú reprezentované zúženiami, ktoré sa nazývajú sekundárne zúženia. Pri elektronickom štúdiu sa rozlišujú fázy vláknitého a granulačného pôvodu.
Vývoj jadra
Ďalšie označenie je fibrilárne, pochádza z proteínu a obrovských polymérov – predchádzajúcich verzií r-RNA. Následne tvoria menšie prvky zrelej rRNA. Keď fibrila dozrieva, stáva sa zrnitou štruktúrou alebo ribonukleoproteínovou granulou.
Chromatín obsiahnutý v štruktúre má farbiace vlastnosti. Je prítomný v nukleoplazme jadra a slúži ako forma medzifázy pre životne dôležitú aktivitu chromozómov. Zloženie chromatínu sú reťazce DNA a polyméry. Spolu tvoria komplex nukleoproteínov.
Históny vykonávajú funkcie organizácie priestoru v štruktúre molekuly DNA. Okrem toho chromozómy zahŕňajú organické látky, enzýmy obsahujúce polysacharidy a kovové častice. Chromatín sa delí na:
- euchromatín,
- heterochromatín.
Prvý je spôsobený nízkou hustotou, takže z takýchto eukaryotov nie je možné prečítať genetické údaje.
Druhá možnosť má kompaktné vlastnosti.
Štruktúra
Konštitúcia samotnej škrupiny je heterogénna. Vďaka neustálym pohybom sa na ňom objavujú výrastky a vydutia. Vo vnútri je to spôsobené pohybmi makromolekúl a ich výstupom do inej vrstvy.
Samotné látky vstupujú 2 spôsobmi:
- fagocytóza,
- pinocytóza.
Fagocytóza sa prejavuje invagináciou pevných častíc. Výčnelky sa nazývajú pinocytóza. Vysunutím sa okraje oblastí približujú k sebe a zachytávajú tekutinu medzi eukaryotmi.
Pinocytóza poskytuje mechanizmus na prenikanie zlúčenín do membrány. Priemer vakuoly sa pohybuje od 0,01 do 1,3 um. Ďalej sa vakuola začne ponoriť do cytoplazmatickej vrstvy a zošnurovať sa. Spojenie medzi bublinami hrá úlohu transportu užitočných častíc a rozkladu enzýmov.
Tráviaci cyklus
Celý okruh tráviacich funkcií je rozdelený do nasledujúcich etáp:
- vstup zložiek do tela,
- rozklad enzýmov
- vstup do cytoplazmy
- vylučovanie.
Prvá fáza zahŕňa vstup látok do ľudského tela. Potom sa začnú rozkladať pomocou lyzozómov. Oddelené častice prenikajú do cytoplazmatického poľa. Nestrávené zvyšky jednoducho vychádzajú von. Následne sa sínus stáva hustým a začína sa premieňať na granulované granule.
Membránové funkcie
Aké funkcie teda plní?
Hlavné budú:
- ochranný,
- prenosný,
- mechanický,
- matica,
- prenos energie,
- receptor.
Ochrana je vyjadrená ako bariéra medzi podjednotkou a vonkajším prostredím. Film slúži ako regulátor výmeny medzi nimi. V dôsledku toho môže byť ten druhý aktívny alebo pasívny. Nastáva selektivita potrebných látok.
V transportnej funkcii sa spojenia prenášajú z jedného mechanizmu do druhého cez plášť. Práve tento faktor ovplyvňuje dodávanie užitočných zlúčenín, odstraňovanie produktov metabolizmu a rozkladu a sekrečných zložiek. Vyvíjajú sa gradienty iónovej povahy, vďaka čomu sa udržiava pH a úroveň koncentrácie iónov.
Posledné dve misie sú pomocné. Práca na úrovni matrice je zameraná na správne umiestnenie proteínového reťazca vo vnútri dutiny a ich správne fungovanie. Vďaka mechanickej fáze je článok zabezpečený v autonómnom režime.
K prenosu energie dochádza v dôsledku fotosyntézy v zelených plastidoch a respiračných procesov v bunkách vo vnútri dutiny. Do práce sa zapájajú aj bielkoviny. Vďaka svojej prítomnosti v membráne poskytujú proteíny makrobunke schopnosť vnímať signály. Impulzy sa pohybujú z jednej cieľovej bunky do ostatných.
Medzi špeciálne vlastnosti membrány patrí generovanie a implementácia biopotenciálu, rozpoznávanie buniek, a teda značenie.
Cytoplazmatická membrána oddeľujúca cytoplazmu od bunkovej steny sa nazýva plazmalema (plazmatická membrána) a oddeľujúca ju od vakuoly sa nazýva tonoplast (elementárna membrána).
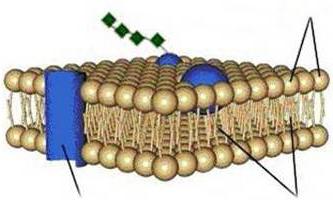
V súčasnosti používajú tekutý mozaikový model membrány (obr. 1.9), podľa ktorého membránu tvorí dvojvrstva lipidových molekúl (fosfolipidov) s hydrofilnými hlavami a 2 hydrofóbnymi chvostíkmi smerujúcimi dovnútra vrstvy. Okrem lipidov obsahujú membrány aj proteíny.
V bilipidovej vrstve „plávajú“ 3 typy membránových proteínov: integrálne proteíny, ktoré prenikajú cez celú hrúbku dvojvrstvy; polointegrálne, neúplne prenikajúce do dvojvrstvy; periférne, pripojené z vonkajšej alebo vnútornej strany membrány k iným membránovým proteínom. Membránové proteíny plnia rôzne funkcie: niektoré z nich sú enzýmy, iné pôsobia ako nosiče špecifických molekúl cez membránu alebo vytvárajú hydrofilné póry, cez ktoré môžu prechádzať polárne molekuly.
Jednou z hlavných vlastností bunkových membrán je ich polopriepustnosť: prepúšťajú vodu, ale neprepúšťajú látky v nej rozpustené, t.j. majú selektívnu priepustnosť.
Ryža. 1.9. Schéma štruktúry biologickej membrány:
A - extracelulárny priestor; B - cytoplazma; 1 - bimolekulárna vrstva lipidov; 2 - periférny proteín; 3 - hydrofilná oblasť integrálneho proteínu; 4 - hydrofóbna oblasť integrálneho proteínu; 5 - sacharidový reťazec
Transport cez membrány
V závislosti od energetického výdaja sa transport látok a iónov cez membránu delí na pasívny, ktorý nevyžaduje energiu, a aktívny, spojený so spotrebou energie. Pasívny transport zahŕňa procesy ako difúzia, uľahčená difúzia a osmóza.
Difúzia je proces prenikania molekúl cez lipidovú dvojvrstvu pozdĺž koncentračného gradientu (z oblasti s vyššou koncentráciou do oblasti s nižšou). Čím je molekula menšia a nepolárnejšia, tým rýchlejšie difunduje cez membránu.
S uľahčenou difúziou pomáha určitý transportný proteín prechodu látky cez membránu. Do bunky tak vstupujú rôzne polárne molekuly, ako sú cukry, aminokyseliny, nukleotidy atď.
Osmóza je difúzia vody cez polopriepustné membrány. Osmóza spôsobuje pohyb vody z roztoku s vysokým vodným potenciálom do roztoku s nízkym vodným potenciálom.
Aktívna doprava- ide o prenos molekúl a iónov cez membránu sprevádzaný nákladmi na energiu. Aktívny transport ide proti koncentračnému gradientu a elektrochemickému gradientu a využíva energiu ATP. Mechanizmus aktívneho transportu látok je založený na práci protónovej pumpy (H+ a K+) v rastlinách a hubách, ktoré udržujú vo vnútri bunky vysokú koncentráciu K+ a nízku koncentráciu H+ (u živočíchov Na+ a K+). Energia potrebná na prevádzku tejto pumpy sa dodáva vo forme ATP, syntetizovaného počas bunkového dýchania.
Je známy ďalší typ aktívneho transportu - endo- a exocytóza. Ide o 2 aktívne procesy, ktorými sú rôzne molekuly transportované cez membránu do bunky ( endocytóza) alebo z neho ( exocytóza).
Pri endocytóze sa látky dostávajú do bunky v dôsledku invaginácie (invaginácie) plazmatickej membrány. Výsledné vezikuly alebo vakuoly sú transportované do cytoplazmy spolu s látkami v nich obsiahnutými. Absorpcia veľkých častíc, ako sú mikroorganizmy alebo zvyšky buniek, sa nazýva fagocytóza. V tomto prípade sa vytvárajú veľké bubliny nazývané vakuoly. Absorpcia kvapalín (suspenzie, koloidné roztoky) alebo rozpustených látok pomocou malých bubliniek je tzv. pinocytóza.
Reverzný proces endocytózy sa nazýva exocytóza. Mnohé látky sa z bunky odstraňujú v špeciálnych vezikulách alebo vakuolách. Príkladom je stiahnutie ich tekutých sekrétov zo sekrečných buniek; ďalším príkladom je účasť diktyozómových vezikúl na tvorbe bunkovej steny.
PROTOPLASTOVÉ DERIVÁTY
Vákuola
Vákuola je rezervoár ohraničený jednou membránou - tonoplastom. Vakuola obsahuje bunkovú šťavu - koncentrovaný roztok rôznych látok, ako sú minerálne soli, cukry, pigmenty, organické kyseliny, enzýmy. V zrelých bunkách sa vakuoly spájajú do jednej, centrálnej.
Vakuoly uchovávajú rôzne látky vrátane konečných produktov metabolizmu. Osmotické vlastnosti bunky silne závisia od obsahu vakuoly.
Vzhľadom na to, že vakuoly obsahujú silné roztoky solí a iných látok, rastlinné bunky neustále osmoticky absorbujú vodu a vytvárajú hydrostatický tlak na bunkovú stenu, nazývaný turgorový tlak. Turgorovmu tlaku odporuje rovnaký tlak z bunkovej steny, nasmerovaný do bunky. Väčšina rastlinných buniek existuje v hypotonickom prostredí. Ale ak sa takáto bunka umiestni do hypertonického roztoku, voda začne bunku opúšťať podľa zákonov osmózy (aby sa vyrovnal potenciál vody na oboch stranách membrány). Vakuola sa zmenší, jej tlak na protoplast sa zníži a membrána sa začne oddeľovať od bunkovej steny. Fenomén oddeľovania protoplastov od bunkovej steny sa nazýva plazmolýza. V prirodzených podmienkach takáto strata turgoru v bunkách povedie k uschnutiu rastliny, ovisnutiu listov a stoniek. Tento proces je však reverzibilný: ak sa bunka vloží do vody (napríklad pri polievaní rastliny), dochádza k javu, ktorý je opakom plazmolýzy – deplazmolýze (pozri obr. 1.10).
Ryža. 1.10. Schéma plazmolýzy:
A - bunka v stave turgoru (v izotonickom roztoku); B - začiatok plazmolýzy (bunka umiestnená v 6% roztoku KNO3); B - úplná plazmolýza (bunka umiestnená v 10% roztoku KNO3); 1 - chloroplast; 2 - jadro; 3 - bunková stena; 4 - protoplast; 5 - centrálna vakuola
Inklúzie
Bunkové inklúzie sú zásobné a vylučovacie látky.
Rezervné látky (dočasne vylúčené z metabolizmu) a s nimi odpadové (vylučovacie látky) sa často nazývajú ergastické látky bunky. Zásobné látky zahŕňajú zásobné bielkoviny, tuky a sacharidy. Tieto látky sa počas vegetácie hromadia v semenách, plodoch, podzemných rastlinných orgánoch a v jadre stonky.
Náhradné látky
Zásobné bielkoviny, príbuzné jednoduchým bielkovinám – bielkovinám, sa často ukladajú v semenách. Vyzrážané proteíny vo vakuolách tvoria okrúhle alebo eliptické zrná nazývané aleurón. Ak aleurónové zrná nemajú rozoznateľnú vnútornú štruktúru a sú zložené z amorfného proteínu, nazývajú sa jednoduché. Ak sa v aleurónových zrnách nachádza medzi amorfným proteínom kryštálovitá štruktúra (kryštaloid) a lesklé bezfarebné telieska okrúhleho tvaru (globoidy), takéto aleurónové zrná sa nazývajú komplexné (pozri obr. 1.11). Amorfný proteín zrna aleurónu je homogénny, nepriehľadný, žltkastý proteín, ktorý napučiava vo vode. Kryštaloidy majú charakteristický romboedrický tvar kryštálov, ale na rozdiel od pravých kryštálov ich bielkovina napučiava vo vode. Globoidy pozostávajú z vápenato-horečnatej soli, obsahujú fosfor, sú nerozpustné vo vode a nereagujú s bielkovinami.
Ryža. 1.11. Komplexné aleurónové zrná:
1 - póry v škrupine; 2 - globoidy; 3 - amorfná proteínová hmota; 4 - kryštaloidy ponorené do amforovej proteínovej hmoty
Zásobné lipidy zvyčajne sa nachádzajú v hyaloplazme vo forme kvapiek a nachádzajú sa takmer vo všetkých rastlinných bunkách. Toto je hlavný typ rezervných živín vo väčšine rastlín: semená a plody sú na ne najbohatšie. Tuky (lipidy) sú najviac kalorickou rezervnou látkou. Činidlom pre látky podobné tuku je Sudan III, ktorý ich farbí na oranžovo.
Sacharidy sú obsiahnuté v zložení každej bunky vo forme vo vode rozpustných cukrov (glukóza, fruktóza, sacharóza) a vo vode nerozpustných polysacharidov (celulóza, škrob). V bunke hrajú sacharidy úlohu zdroja energie pre metabolické reakcie. Cukry po naviazaní s inými biologickými látkami bunky tvoria glykozidy a polysacharidy s bielkovinami glykoproteíny. Zloženie uhľohydrátov v rastlinnej bunke je oveľa rozmanitejšie ako v živočíšnych bunkách, čo je spôsobené rôznorodým zložením polysacharidov bunkovej steny a cukrov v bunkovej šťave vakuol.
Hlavným a najbežnejším zásobným sacharidom je polysacharidový škrob. Primárny asimilačný škrob sa tvorí v chloroplastoch. V noci, keď sa fotosyntéza zastaví, sa škrob hydrolyzuje na cukry a transportuje do zásobných tkanív – hľúz, cibúľ, podzemkov. Tam sa v špeciálnych typoch leukoplastov – amyloplastoch – časť cukrov ukladá vo forme zŕn sekundárneho škrobu. Škrobové zrná sa vyznačujú vrstvením, čo sa vysvetľuje rozdielnym obsahom vody v dôsledku nerovnomerného prísunu škrobu počas dňa. V tmavých vrstvách je viac vody ako vo svetlých vrstvách. Zrno s jedným stredom tvorby škrobu v strede amyloplastu sa nazýva jednoduché koncentrické, ak je stred posunuté, nazýva sa jednoduché excentrické. Zrno s niekoľkými centrami tvoriacimi škrob je zložité. V polozložených zrnách sa okolo niekoľkých škrobotvorných centier uložia nové vrstvy a potom sa vytvoria spoločné vrstvy a pokryjú škrobotvorné centrá (pozri obr. 1.12). Činidlom pre škrob je roztok jódu, ktorý dáva modrú farbu.
Ryža. 1.12. Zrná zemiakového škrobu (A):
1 - jednoduché zrno; 2 - polokomplex; 3 - komplex; pšenica (B), ovos (C)
Vylučovacie látky (sekundárne metabolické produkty)
Bunkové inklúzie zahŕňajú aj vylučovacie látky, napríklad kryštály šťavelanu vápenatého ( monokryštály, raphidy - ihličkovité kryštály, drúzy - kryštálové zrasty, kryštalický piesok - nahromadenie mnohých malých kryštálikov) (pozri obr. 1.13). Menej často sa kryštály skladajú z uhličitanu vápenatého alebo oxidu kremičitého ( cystolity; pozri obr. 1.14). Cystolity sú uložené na bunkovej stene, vyčnievajúce do bunky v podobe strapcov hrozna a sú charakteristické napríklad pre zástupcov čeľade žihľavy a listy fikusu.
Na rozdiel od zvierat, ktoré prebytočné soli vylučujú močom, rastliny nemajú vyvinuté vylučovacie orgány. Preto sa predpokladá, že kryštály šťavelanu vápenatého sú konečným produktom metabolizmu protoplastov, vytvoreným ako zariadenie na odstraňovanie nadbytočného vápnika z metabolizmu. Tieto kryštály sa spravidla hromadia v orgánoch, ktoré rastlina pravidelne vylučuje (listy, kôra).

Ryža. 1.13. Formy kryštálov šťavelanu vápenatého v bunkách:
1, 2 - raphida (impatiens; 1 - bočný pohľad, 2 - rez); 3 - drúza (opuncia); 4 - kryštalický piesok (zemiaky); 5 - monokryštál (vanilka)
Ryža. 1.14. Cystolit (na priereze listu fikusu):
1 - šupka listov; 2 - cystolitída
Esenciálne oleje hromadia sa v listoch (mäta, levanduľa, šalvia), kvetoch (šípky), plodoch (citrusové plody) a semenách rastlín (kôpor, aníz). Éterické oleje sa nezúčastňujú metabolizmu, ale sú široko používané v parfumérii (ružové, jazmínové oleje), potravinárstve (aníz, kôprové oleje), medicíne (mäta, eukalyptové oleje). Zásobníky na akumuláciu éterických olejov môžu byť žľazy (mäta), lyzogénne nádoby (citrusové plody), žľazové chĺpky (pelargónie).
Živice- ide o komplexné zlúčeniny vznikajúce počas normálneho života alebo v dôsledku deštrukcie tkaniva. Sú tvorené epiteliálnymi bunkami, ktoré lemujú živicové kanáliky ako vedľajší produkt metabolizmu, často s éterickými olejmi. Môžu sa hromadiť v bunkovej šťave, cytoplazme vo forme kvapiek alebo v nádobách. Sú nerozpustné vo vode, nepriepustné pre mikroorganizmy a vďaka svojim antiseptickým vlastnostiam zvyšujú odolnosť rastlín voči chorobám. Živice sa používajú v medicíne, ako aj pri výrobe farieb, lakov a mazacích olejov. V modernom priemysle ich nahrádzajú syntetické materiály.
Bunková stena
Pevná bunková stena obklopujúca bunku pozostáva z celulózových mikrofibríl uložených v matrici obsahujúcej hemicelulózy a pektínové látky. Bunková stena poskytuje mechanickú oporu bunke, chráni protoplast a udržuje tvar bunky. V tomto prípade je bunková stena schopná natiahnutia. Stena, ktorá je produktom životne dôležitej aktivity protoplastu, môže rásť iba v kontakte s ním. Cez bunkovú stenu sa pohybuje voda a minerálne soli, tá je však úplne alebo čiastočne nepriepustná pre vysokomolekulárne látky. Keď protoplast odumrie, stena môže naďalej vykonávať funkciu vedenia vody. Prítomnosť bunkovej steny, viac ako všetky ostatné charakteristiky, odlišuje rastlinné bunky od živočíšnych. Architektúru bunkovej steny do značnej miery určuje celulóza. Monomérom celulózy je glukóza. Zväzky molekúl celulózy tvoria micely, ktoré sa spájajú do väčších zväzkov – mikrofibríl. Činidlom pre celulózu je chlór-zinok-jód (Cl-Zn-I), ktorý dáva modrofialové sfarbenie.
Celulózová štruktúra bunkovej steny je vyplnená molekulami necelulózovej matrice. Matrica obsahuje polysacharidy nazývané hemicelulózy; pektínové látky (pektín), veľmi blízke hemicelulózam, a glykoproteíny. Pektické látky, ktoré sa spájajú medzi susednými bunkami, vytvárajú strednú platňu, ktorá sa nachádza medzi primárnymi membránami susedných buniek. Keď sa stredná platňa rozpustí alebo zničí (čo sa vyskytuje v dužine dozretých plodov), dochádza k macerácii (z latinského maceratio - zmäkčenie). Prirodzenú maceráciu možno pozorovať u mnohých prezretých plodov (melón, melón, broskyňa). Umelá macerácia (keď je tkanivo ošetrené zásadou alebo kyselinou) sa používa na prípravu rôznych anatomických a histologických preparátov.
Bunková stena v procese života môže prechádzať rôznymi úpravami - lignifikáciou, suberizáciou, slizom, kutinizáciou, mineralizáciou (pozri tabuľku l.4).
Tabuľka 1.4.
Súvisiace informácie.