
Ce proprietăți are membrana citoplasmatică? Membrana celulara
Membrana citoplasmatică exterioară este o peliculă subțire. Grosimea sa este de aproximativ 7-10 nm. Filmul este vizualizat doar la microscop electronic.
Structura
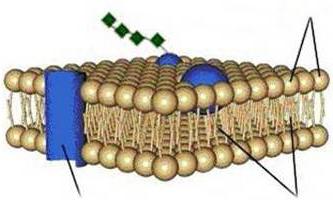
Care este compoziția membranei citoplasmatice? Structura filmului este destul de diversă. Conform organizării sale chimice, este un complex de proteine și lipide. Membrana citoplasmatică a unei celule include un strat dublu. Acționează ca bază. În plus, membrana citoplasmatică conține colesterol și glicolipide. Aceste substanțe se caracterizează prin amfipatricitate. Cu alte cuvinte, ele conțin capete hidrofobe („frică de apă”) și hidrofile („iubitoare de apă”). Acestea din urmă (grupa fosfat) sunt direcționate spre exterior din membrană, cele din urmă (reziduuri din acizi grași) sunt orientate unul spre celălalt. Din acest motiv, se formează un strat bipolar lipidic. Moleculele de lipide au mobilitate. Ei sunt capabili să se deplaseze în propriul lor monostrat sau (rar) de la unul la altul.

Stratul lipidic poate fi în stare solidă sau lichidă. Monostraturile sunt asimetrice. Aceasta înseamnă că compoziția lor lipide este diferită. Datorită acestei proprietăți, membranele citoplasmatice au specificitate chiar și în cadrul unei singure celule. A doua componentă esențială a filmului include proteine. Multe dintre aceste conexiuni se pot deplasa în planul membranei sau se pot roti în jurul propriei axe. Cu toate acestea, ei nu sunt capabili să se deplaseze dintr-o parte a stratului dublu în alta. Protejarea mediului intern este sarcina principală îndeplinită de membrana citoplasmatică. Structura filmului, în plus, asigură cursul diferitelor procese. Proteinele sunt responsabile pentru îndeplinirea anumitor sarcini. Lipidele oferă caracteristicile structurale ale filmului.
Membrana citoplasmatica: functii
Sarcinile principale sunt:
- Barieră. Filmul protector asigură schimbul activ, pasiv, selectiv, reglat de compuși cu mediul extern. Datorită permeabilității selective, celula și compartimentele sale sunt separate și alimentate cu substanțele necesare.
- Transport. Tranziția compușilor de la celulă la celulă are loc prin film. Datorită acestui fapt, sunt eliberați compuși nutritivi, produsele finale metabolice sunt îndepărtate și sunt secretate diferite substanțe. În plus, se formează gradienți ionici și concentrația ionică și pH-ul sunt menținute la niveluri optime. Sunt necesare pentru activitatea activă a enzimelor celulare.
Sarcini auxiliare

Proprietăți speciale
Funcțiile specifice ale membranei includ:

Informații suplimentare
Dacă unele particule, dintr-un motiv sau altul, nu sunt capabile să treacă prin stratul dublu fosfolipidic (de exemplu, datorită proprietăților hidrofile, deoarece membrana citoplasmatică din interior este hidrofobă și nu permite trecerea unor astfel de compuși sau datorită dimensiunea particulelor în sine), dar sunt necesare, apoi pot trece cu ajutorul proteinelor purtătoare speciale (transportatori) și proteinelor canalului. Sau pătrunderea lor se realizează prin endocitoză.
În procesul de transport pasiv, substanțele traversează stratul lipidic prin difuzie. În acest caz, nu se irosește energie. Difuzia facilitată poate fi una dintre variantele unui astfel de mecanism. În timpul acestui proces, o moleculă specifică facilitează trecerea substanței. Poate avea un canal capabil să treacă numai particule de același tip. Transportul activ consumă energie. Acest lucru se datorează faptului că acest proces se efectuează împotriva unui gradient de concentrație. Membrana citoplasmatică conține proteine speciale de pompă, inclusiv ATPaza, care favorizează intrarea activă a ionilor de potasiu și excreția ionilor de sodiu.
Modele
Există mai multe dintre ele:
- "Model sandwich". Ideea unei structuri cu trei straturi a tuturor membranelor a fost exprimată de oamenii de știință Dawson și Danieli în 1935. În opinia lor, structura filmului era următoarea: proteine-lipide-proteine. Această idee a existat de destul de mult timp.
- „Structură lichid-mozaic”. Acest model a fost descris de Nicholson și Singer în 1972. În conformitate cu acesta, moleculele de proteine nu formează un strat continuu, ci sunt scufundate într-un strat lipidic bipolar sub forma unui mozaic la adâncimi diferite. Acest model este considerat cel mai universal.
- „Structura cristalină a proteinei”. Conform acestui model, membranele se formează datorită împleterii moleculelor de proteine și lipide, care sunt combinate pe baza legăturilor hidrofil-hidrofobe.
Orice celulă vie este separată de mediu printr-o membrană subțire cu o structură specială - membrana citoplasmatică (CPM). Eucariotele au numeroase membrane intracelulare care separă spațiul organelelor de citoplasmă, în timp ce pentru majoritatea procariotelor CPM este singura membrană celulară. În unele bacterii și arhei, poate pătrunde în citoplasmă, formând excrescențe și pliuri de diferite forme.
CPM-urile oricăror celule sunt construite conform unui singur plan și constau din fosfolipide (Fig. 3.5, A). La bacterii, ei conțin doi acizi grași, de obicei cu 16-18 atomi de carbon în lanț și cu legături saturate sau nesaturate, conectați printr-o legătură esterică la două grupări hidroxil ale glicerolului. Compoziția de acizi grași a bacteriilor poate varia ca răspuns la schimbările de mediu, în special temperatura. Când temperatura scade, cantitatea de acizi grași nesaturați din compoziția fosfolipidelor crește, ceea ce afectează semnificativ fluiditatea membranei. Unii acizi grași pot fi ramificați sau pot conține un inel ciclopropan. A treia grupă OH a glicerolului este conectată la reziduul de acid fosforic și prin acesta la grupul principal. Grupurile principale ale fosfolipidelor pot avea naturi chimice diferite la diferite procariote (fosfatidiletanolamină, fosfatidilglicerol, cardiolipină, fosfatidilserina, lecitină etc.), dar sunt mai simple ca structură decât la eucariote. De exemplu, la E coli, sunt reprezentați de 75% fosfatidiletanolamină, 20% fosfatidilglicerol, restul constau din cardiolipină (difosfatidilglicerol), fosfatidilserina și urme de alți compuși. Alte bacterii au tipuri mai complexe de lipide membranare. Unele celule formează glicolipide, cum ar fi monogalactosil diglicerida. Lipidele membranei arheale diferă de cele eucariote și bacteriene. În loc de acizi grași, ei conțin alcooli izoprenoizi mai mari atașați la glicerol printr-o legătură simplă, mai degrabă decât ester.
Orez. 3.5.
A- fosfolipide; b- membrana dublu stratificata
O O o O o o
Astfel de molecule alcătuiesc un dublu strat de membrană, unde părțile hidrofobe sunt orientate spre interior, iar părțile hidrofile spre exterior, în mediu și în citoplasmă (Fig. 3.5, b). Numeroase proteine sunt încorporate în stratul dublu sau se intersectează și pot difuza în interiorul membranei, formând uneori complexe complexe. Proteinele membranare au o serie de funcții importante, inclusiv conversia și stocarea energiei metabolice, reglarea absorbției și eliberării tuturor nutrienților și produselor metabolice. În plus, ei recunosc și transmit multe semnale care reflectă schimbările din mediu și declanșează cascada corespunzătoare de reacții care conduc la un răspuns celular. Această organizare a membranelor este bine explicată prin modelul cu cristale lichide cu un mozaic intercalate cu proteine membranare (Fig. 3.6).

Orez. 3.6.
Majoritatea membranelor biologice au o grosime de 4 până la 7 nm. Membranele celulare sunt clar vizibile într-un microscop electronic cu transmisie atunci când sunt în contrast cu metalele grele. În micrografiile electronice ele arată ca formațiuni cu trei straturi: două straturi exterioare întunecate arată poziția grupurilor polare de lipide, iar stratul mediu luminos arată spațiul interior hidrofob (Fig. 3.7).
O altă tehnică pentru studierea membranelor este obținerea de celule scindate congelate la temperatura azotului lichid și contrastarea suprafețelor rezultate prin pulverizarea metalelor grele.
(platină, aur, argint). Preparatele rezultate sunt vizualizate la un microscop electronic cu scanare. În acest caz, se poate vedea suprafața membranei și proteinele membranei mozaic incluse în aceasta, care nu se extind prin membrană, ci sunt conectate prin regiuni speciale de ancorare hidrofobe de regiunea hidrofobă a stratului dublu.

Orez. 3.7.
CPM are proprietatea de permeabilitate selectivă, împiedicând mișcarea liberă a majorității substanțelor în și în afara celulei și, de asemenea, joacă un rol semnificativ în creșterea și diviziunea celulară, mișcarea și exportul proteinelor de suprafață și extracelulare și carbohidraților (exopolizaharide). . Dacă o celulă este plasată într-un mediu cu o presiune osmotică mai mare sau mai mică decât în interiorul citoplasmei, atunci apa va părăsi celula sau apa va intra în ea. Aceasta reflectă proprietatea apei de a egaliza gradienții de soluție. In acest caz, citoplasma se contracta sau se extinde (fenomenul de plasmoliza/deplasmoliza). Majoritatea bacteriilor, cu toate acestea, nu își schimbă forma în astfel de experimente din cauza prezenței unui perete celular rigid.
CPM reglează fluxul de nutrienți și metaboliți. Prezența unui strat hidrofob format din lipide membranare împiedică trecerea oricăror molecule polare și macromolecule prin acesta. Această proprietate permite celulelor, care există în general în soluții diluate, să rețină macromolecule utile și precursori metabolici. Membrana celulară este, de asemenea, proiectată pentru a îndeplini o funcție de transport. De obicei, procariotele au un număr mare de sisteme de transport foarte specifice. Transportul este o parte integrantă a bioenergeticii globale a celulei, care creează și utilizează diverși gradienți ionici prin CPM pentru a transporta substanțe și a forma alți gradienți necesari celulei. CPM joacă un rol semnificativ în mișcarea, creșterea și diviziunea celulelor. Multe procese metabolice sunt concentrate în membrana procariotelor. Proteinele membranare îndeplinesc funcții importante: participă la transformarea și stocarea energiei, reglează absorbția și eliberarea tuturor nutrienților și produselor metabolice, recunosc și transmit semnale despre schimbările din mediu.
Membrana citoplasmatică exterioară care înconjoară citoplasma fiecărei celule determină dimensiunea acesteia și asigură menținerea unor diferențe semnificative între conținutul celular și mediu. Membrana servește ca un filtru foarte selectiv care menține o diferență în concentrațiile ionilor pe ambele părți ale membranei și permite nutrienților să pătrundă în celulă și deșeurilor să părăsească celula.
Toate membranele biologice sunt ansambluri de molecule de lipide și proteine ținute împreună prin interacțiuni non-covalente. Moleculele de lipide și proteine formează un strat dublu continuu.
Bistratul lipidic este structura principală a membranei, care creează o barieră relativ impermeabilă pentru majoritatea moleculelor solubile în apă.
Moleculele de proteine sunt, parcă, „dizolvate” în stratul dublu lipidic. Prin intermediul proteinelor se realizează diverse funcții membranare: unele dintre ele asigură transportul anumitor molecule în sau în afara celulei, altele sunt enzime și catalizează reacții asociate membranei, iar altele asigură o legătură structurală între citoschelet și matricea extracelulară sau servesc. ca receptori pentru recepția și convertirea semnalelor chimice din mediu.
O proprietate importantă a membranelor biologice este fluiditatea. Toate membranele celulare sunt structuri fluide mobile: majoritatea moleculelor lor constitutive de lipide și proteine sunt capabile să se miște destul de repede în planul membranei. O altă proprietate a membranelor este asimetria lor: ambele straturi diferă în ceea ce privește compoziția de lipide și proteine, ceea ce reflectă diferențele funcționale ale suprafețelor lor.
Funcțiile membranei citoplasmatice exterioare:
· bariera – asigura metabolismul reglat, selectiv, pasiv si activ cu mediul. Permeabilitatea selectivă asigură că celulele și compartimentele celulare sunt separate de mediu și aprovizionate cu substanțele necesare.
Transport - transportul substanțelor în și din celulă are loc prin membrană. Transportul prin membrane asigură: livrarea nutrienților, îndepărtarea produselor finite metabolice, secreția diferitelor substanțe, crearea gradienților de ioni, menținerea pH-ului și concentrației ionice adecvate în celulă, care sunt necesare pentru funcționarea enzimelor celulare.
Particule care din anumite motive nu sunt capabile să traverseze stratul dublu fosfolipidic (de exemplu, datorită proprietăților hidrofile, deoarece membrana din interior este hidrofobă și nu permite trecerea substanțelor hidrofile sau datorită dimensiunilor lor mari), dar necesare pentru celula, poate patrunde in membrana prin proteine transportoare speciale (transportatori) si proteine canale sau prin endocitoza.
În timpul transportului pasiv, substanțele traversează bistratul lipidic fără consum de energie, prin difuzie. O variantă a acestui mecanism este difuzia facilitată, în care o moleculă specifică ajută o substanță să treacă prin membrană. Această moleculă poate avea un canal care permite trecerea unui singur tip de substanță.
Transportul activ necesită energie, deoarece are loc împotriva unui gradient de concentrație. Există proteine speciale de pompă pe membrană, inclusiv ATPaza, care pompează activ ionii de potasiu (K+) în celulă și pompează ionii de sodiu (Na+) din ea.
· matrice - asigură o anumită poziție relativă și orientare a proteinelor membranare, interacțiunea optimă a acestora;
· mecanic - asigură autonomia celulei, structurile ei intracelulare, precum și legătura cu alte celule (în țesuturi). Pereții celulari joacă un rol major în asigurarea funcției mecanice, iar la animale, substanța intercelulară.
· energie - în timpul fotosintezei în cloroplaste și a respirației celulare în mitocondrii, în membranele lor funcționează sisteme de transfer de energie, la care participă și proteinele;
· receptor - unele proteine care se află în membrană sunt receptori (molecule cu ajutorul cărora celula percepe anumite semnale).
De exemplu, hormonii care circulă în sânge acționează doar asupra celulelor țintă care au receptori corespunzători acestor hormoni. Neurotransmițătorii (substanțe chimice care asigură conducerea impulsurilor nervoase) se leagă și de proteinele receptorilor speciale din celulele țintă.
· enzimatice - proteinele membranare sunt adesea enzime. De exemplu, membranele plasmatice ale celulelor epiteliale intestinale conțin enzime digestive.
· implementarea generarii si conducerii biopotentialelor.
Cu ajutorul membranei, în celulă se menține o concentrație constantă de ioni: concentrația ionului K+ în interiorul celulei este mult mai mare decât în exterior, iar concentrația Na+ este mult mai mică, ceea ce este foarte important, deoarece acest lucru asigură menținerea diferenței de potențial pe membrană și generarea unui impuls nervos.
· marcarea celulelor – există antigene pe membrană care acționează ca markeri – „etichete” care permit identificarea celulei. Acestea sunt glicoproteine (adică proteine cu lanțuri laterale de oligozaharide ramificate atașate la acestea) care joacă rolul de „antene”. Datorită multitudinii de configurații ale lanțurilor laterale, este posibil să se facă un marker specific pentru fiecare tip de celulă. Cu ajutorul markerilor, celulele pot recunoaște alte celule și pot acționa împreună cu acestea, de exemplu, în formarea organelor și țesuturilor. Acest lucru permite, de asemenea, sistemului imunitar să recunoască antigenele străine.
Fiecare corp uman sau animal este format din miliarde de celule. O celulă este un mecanism complex care îndeplinește funcții specifice. Toate organele și țesuturile sunt formate din subunități.
Sistemul are o membrană citoplasmatică, citoplasmă, nucleu și o serie de organite. Nucleul este separat de organele printr-o membrană internă. Toate împreună oferă viață țesuturilor și, de asemenea, permit metabolismul.
Lema plasmatică citoplasmatică sau membrana joacă un rol important în funcționare.
Numele în sine, membrana citoplasmatică exterioară, provine din latină membrana, sau altfel piele. Acesta este un delimitator de spațiu între organismele celulare.
Ipoteza structurii a fost înaintată deja în 1935. În 1959, V. Robertson a ajuns la concluzia că învelișurile membranelor sunt aranjate după același principiu.
Datorită cantității mari de informații acumulate, cavitatea a dobândit un model mozaic lichid al structurii. Acum este considerat universal acceptat. Este membrana citoplasmatică exterioară care formează învelișul exterior al unităților.
Structura
Deci, ce este lema plasmatică?
Este o peliculă subțire care separă procariotele de mediul intern. Poate fi văzut doar la microscop. Structura membranei citoplasmatice include un strat dublu, care servește ca bază.
Bistratul este un strat dublu format din proteine și lipide. Există, de asemenea, colesterolul și glicolipidele, care sunt amfipatrice.
Ce înseamnă?
Organismul gras are un cap bipolar și o coadă hidrofilă. Primul se datorează fricii de apă, iar al doilea se datorează absorbției acesteia. Grupul de fosfați are o direcție spre exterior din film, cei din urmă sunt îndreptați unul spre celălalt.
Astfel, se formează un strat lipidic bipolar. Lipidele sunt foarte active, se pot deplasa în monostratul lor și rareori se deplasează în alte zone.
Polimerii sunt împărțiți în:
- extern,
- integral,
- pătrunzând lema plasmatică.
Primele sunt localizate doar pe partea superficială a sinusului. Ele sunt ținute împreună prin electrostatică cu capetele bipolare ale elementelor lipidice. Reține enzimele nutritive. Integrale în interior, sunt încorporate în structura cochiliei în sine, conexiunile își schimbă locația datorită mișcării eucariotelor. Acestea servesc ca un fel de transportor, construite astfel încât substraturile și produsele de reacție să curgă de-a lungul lor. Compușii proteici care pătrund în macrocavitatea au proprietățile de a forma pori pentru intrarea nutrienților în organism.
Miez
Orice unitate are un nucleu, aceasta este baza sa. Membrana citoplasmatică are, de asemenea, un organel, a cărui structură va fi descrisă mai jos.
Structura nucleară include membrana, seva, locul de asamblare a ribozomilor și cromatina. Învelișul este împărțit de spațiul nuclear, este înconjurat de lichid.
Funcțiile organelelor sunt împărțite în două principale:
- închiderea structurii în organite,
- reglarea miezului și a conținutului de lichid.
Miezul este format din pori, fiecare determinat de prezența combinațiilor de pori grei. Volumul lor poate indica capacitatea motrică activă a eucariotelor. De exemplu, imaturele cu activitate ridicată conțin un număr mai mare de pori. Proteinele servesc ca suc nuclear.
Polimerii reprezintă o combinație de matrice și nucleoplasmă. Lichidul este conținut în interiorul peliculei nucleare și asigură funcționalitatea conținutului genetic al organismelor. Elementul proteic oferă protecție și rezistență subunităților.
ARN-urile ribozomale se maturizează în nucleol însuși. Genele ARN în sine sunt localizate pe o regiune specifică a mai multor cromozomi. În cadrul lor se formează mici organizatori. Nucleolii înșiși sunt creați în interior. Zonele din cromozomii mitotici sunt reprezentate de constricții, numite constricții secundare. La studierea electronică se disting fazele de origine fibroasă și granulativă.
Dezvoltare de bază
O altă denumire este fibrilară, provine din proteine și polimeri uriași - versiuni anterioare de r-ARN. Ulterior, ele formează elemente mai mici de ARNr matur. Când fibrila se maturizează, devine granulară ca structură sau granulă de ribonucleoproteină.
Cromatina inclusă în structură are proprietăți de colorare. Prezent în nucleoplasma nucleului, servește ca formă de interfază pentru activitatea vitală a cromozomilor. Compoziția cromatinei este catene și polimeri de ADN. Împreună formează un complex de nucleoproteine.
Histonele îndeplinesc funcțiile de organizare a spațiului în structura moleculei de ADN. În plus, cromozomii includ substanțe organice, enzime care conțin polizaharide și particule de metal. Cromatina este împărțită în:
- eucromatina,
- heterocromatina.
Primul se datorează densității scăzute, deci este imposibil să citiți date genetice de la astfel de eucariote.
A doua opțiune are proprietăți compacte.
Structura
Constituția învelișului în sine este eterogenă. Datorită mișcărilor constante, pe ea apar creșteri și umflături. În interior, acest lucru se datorează mișcărilor macromoleculelor și ieșirii lor într-un alt strat.
Substanțele în sine intră în 2 moduri:
- fagocitoză,
- pinocitoza.
Fagocitoza se exprimă prin invaginarea particulelor solide. Bulbiile se numesc pinocitoză. Prin proeminență, marginile regiunilor se apropie, captând fluidul între eucariote.
Pinocitoza oferă un mecanism de penetrare a compușilor în membrană. Diametrul vacuolei variază de la 0,01 la 1,3 um. În continuare, vacuola începe să se scufunde în stratul citoplasmatic și să se încingă. Legătura dintre bule joacă rolul de a transporta particule utile și de a descompune enzimele.
Ciclul digestiv
Întregul cerc al funcției digestive este împărțit în următoarele etape:
- intrarea componentelor în organism,
- defalcarea enzimelor
- intrarea în citoplasmă
- excreţie.
Prima fază presupune intrarea substanțelor în corpul uman. Apoi încep să se descompună cu ajutorul lizozomilor. Particulele separate pătrund în câmpul citoplasmatic. Reziduurile nedigerate ies pur și simplu în mod natural. Ulterior, sinusul devine dens și începe să se transforme în granule granulare.
Funcțiile membranei
Deci, ce funcții îndeplinește?
Principalele vor fi:
- de protecţie,
- portabil,
- mecanic,
- matrice,
- transfer de energie,
- receptor.
Protecția este exprimată ca o barieră între subunitate și mediul extern. Filmul servește ca un regulator al schimbului dintre ei. Ca urmare, acesta din urmă poate fi activ sau pasiv. Apare selectivitatea substanțelor necesare.
În funcția de transport, conexiunile sunt transferate de la un mecanism la altul prin intermediul carcasei. Acest factor afectează livrarea compușilor utili, eliminarea produselor metabolice și de degradare și a componentelor secretoare. Se dezvoltă gradienți de natură ionică, datorită cărora se mențin pH-ul și nivelul concentrației ionilor.
Ultimele două misiuni sunt auxiliare. Lucrările la nivel de matrice vizează localizarea corectă a lanțului proteic în interiorul cavității și buna funcționare a acestora. Datorita fazei mecanice, celula este asigurata in regim autonom.
Transferul de energie are loc ca urmare a fotosintezei în plastidele verzi și a proceselor respiratorii în celulele din interiorul cavității. Proteinele sunt, de asemenea, implicate în lucru. Datorită prezenței lor în membrană, proteinele oferă macrocelulei capacitatea de a percepe semnale. Impulsurile se deplasează de la o celulă țintă la restul.
Proprietățile speciale ale membranei includ generarea și implementarea biopotențialului, recunoașterea celulelor și adică etichetarea.
Membrana citoplasmatică care separă citoplasma de peretele celular se numește plasmalemă (membrană plasmatică), iar separarea acesteia de vacuole se numește tonoplast (membrană elementară).
În prezent, se utilizează modelul mozaic lichid al membranei (Fig. 1.9), conform căruia membrana este formată dintr-un strat dublu de molecule lipidice (fosfolipide) cu capete hidrofile și 2 cozi hidrofobe orientate spre interiorul stratului. Pe lângă lipide, membranele conțin și proteine.
Există 3 tipuri de proteine membranare „plutitoare” în stratul bilipid: proteine integrale care pătrund în toată grosimea stratului dublu; semi-integral, pătrunzând incomplet stratul dublu; periferice, atașate din partea exterioară sau interioară a membranei de alte proteine membranare. Proteinele membranei îndeplinesc diverse funcții: unele dintre ele sunt enzime, altele acționează ca purtători de molecule specifice peste membrană sau formează pori hidrofili prin care pot trece molecule polare.
Una dintre principalele proprietăți ale membranelor celulare este semi-permeabilitatea lor: permit trecerea apei, dar nu permit trecerea substanțelor dizolvate în ea, adică au permeabilitate selectivă.
Orez. 1.9. Schema structurii unei membrane biologice:
A - spațiu extracelular; B - citoplasmă; 1 - stratul bimolecular de lipide; 2 - proteina periferica; 3 - regiunea hidrofilă a proteinei integrale; 4 - regiunea hidrofobă a proteinei integrale; 5 - lanțul de carbohidrați
Transport prin membrane
În funcție de consumul de energie, transportul substanțelor și ionilor prin membrană se împarte în pasiv, care nu necesită energie, și activ, asociat cu consumul de energie. Transportul pasiv include procese precum difuzia, difuzia facilitată și osmoza.
Difuzia este procesul de penetrare a moleculelor printr-un strat dublu lipidic de-a lungul unui gradient de concentrație (de la o zonă de concentrație mai mare la o zonă de mai mică). Cu cât molecula este mai mică și mai nepolară, cu atât difuzează mai repede prin membrană.
Cu difuzie facilitată, unele proteine de transport ajută la trecerea unei substanțe prin membrană. Astfel, în celulă intră diverse molecule polare, precum zaharuri, aminoacizi, nucleotide etc.
Osmoză este difuzia apei prin membranele semipermeabile. Osmoza determină mișcarea apei dintr-o soluție cu potențial de apă ridicat la o soluție cu potențial de apă scăzut.
Transport activ- este transferul de molecule și ioni printr-o membrană, însoțit de costuri energetice. Transportul activ merge împotriva gradientului de concentrație și a gradientului electrochimic și folosește energia ATP. Mecanismul de transport activ al substanțelor se bazează pe activitatea pompei de protoni (H+ și K+) la plante și ciuperci, care mențin o concentrație mare de K+ și o concentrație scăzută de H+ în interiorul celulei (Na+ și K+ la animale). Energia necesară pentru a funcționa această pompă este furnizată sub formă de ATP, sintetizat în timpul respirației celulare.
Un alt tip de transport activ este cunoscut - endo- și exocitoza. Acestea sunt două procese active prin care diferite molecule sunt transportate prin membrană în celulă ( endocitoza) sau din ea ( exocitoza).
În timpul endocitozei, substanțele intră în celulă ca urmare a invaginării (invaginării) membranei plasmatice. Veziculele rezultate, sau vacuolele, sunt transportate în citoplasmă împreună cu substanțele conținute în ele. Absorbția particulelor mari, cum ar fi microorganismele sau resturile celulare, se numește fagocitoză. În acest caz, se formează bule mari numite vacuole. Absorbția lichidelor (suspensii, soluții coloidale) sau a substanțelor dizolvate cu ajutorul bulelor mici se numește pinocitoza.
Procesul invers al endocitozei se numește exocitoză. Multe substanțe sunt îndepărtate din celulă în vezicule sau vacuole speciale. Un exemplu este retragerea secrețiilor lor lichide din celulele secretoare; un alt exemplu este participarea veziculelor de dictiozom la formarea peretelui celular.
DERIVATI PROTOPLAST
Vacuole
Vacuole- Acesta este un rezervor delimitat de o singură membrană - tonoplastul. Vacuola conține seva celulară - o soluție concentrată de diferite substanțe, precum săruri minerale, zaharuri, pigmenți, acizi organici, enzime. În celulele mature, vacuolele se contopesc într-una, centrală.
Vacuolele stochează diverse substanțe, inclusiv produsele finale metabolice. Proprietățile osmotice ale celulei depind puternic de conținutul vacuolei.
Datorită faptului că vacuolele conțin soluții puternice de săruri și alte substanțe, celulele plantelor absorb în mod osmotic apa și creează presiune hidrostatică pe peretele celular, numită presiune de turgescență. Presiunea turgenței este opusă unei presiuni egale din peretele celular, direcționată în celulă. Majoritatea celulelor vegetale există într-un mediu hipotonic. Dar dacă o astfel de celulă este plasată într-o soluție hipertonică, apa va începe să părăsească celula conform legilor osmozei (pentru a egaliza potențialul de apă de pe ambele părți ale membranei). Vacuola se va micșora în volum, presiunea sa asupra protoplastului va scădea, iar membrana va începe să se îndepărteze de peretele celular. Fenomenul de detașare a protoplastelor de peretele celular se numește plasmoliză. În condiții naturale, o astfel de pierdere a turgenței în celule va duce la ofilirea plantei, la căderea frunzelor și a tulpinilor. Cu toate acestea, acest proces este reversibil: dacă o celulă este plasată în apă (de exemplu, la udarea unei plante), are loc un fenomen care este opusul plasmolizei - deplasmoliza (vezi Fig. 1.10).
Orez. 1.10. Schema de plasmoliza:
A - celulă în stare de turgescență (în soluție izotonă); B - începutul plasmolizei (celulă plasată într-o soluție 6% KNO3); B - plasmoliza completă (celulă plasată într-o soluție 10% KNO3); 1 - cloroplast; 2 - miez; 3 - peretele celular; 4 - protoplast; 5 - vacuola centrala
Incluziuni
Incluziunile celulare sunt substanțe de depozitare și excreție.
Substantele de rezerva (excluse temporar din metabolism) si odata cu ele deseurile (substantele excretoare) sunt adesea numite substante ergastice ale celulei. Substanțele de depozitare includ proteine de depozitare, grăsimi și carbohidrați. Aceste substanțe se acumulează în timpul sezonului de vegetație în semințe, fructe, organe subterane ale plantelor și în miezul tulpinii.
Substanțe de rezervă
Proteinele de depozitare, legate de proteinele simple – proteine, sunt deseori depuse în semințe. Proteinele precipitate în vacuole formează boabe rotunde sau eliptice numite aleuronă. Dacă boabele de aleuronă nu au o structură internă vizibilă și sunt compuse din proteine amorfe, se numesc simple. Dacă în boabele de aleuronă o structură asemănătoare cristalului (cristaloid) și corpuri strălucitoare, incolore, de formă rotundă (globoide) se găsesc printre proteinele amorfe, astfel de boabe de aleuronă se numesc complexe (vezi Fig. 1.11). Proteina amorfa a boabelor de aleurona este o proteina omogena, opaca, galbuie, care se umfla in apa. Cristaloizii au forma romboedrică caracteristică a cristalelor, dar spre deosebire de cristalele adevărate, proteina lor constitutivă se umflă în apă. Globoizii constau dintr-o sare de calciu-magneziu, contin fosfor, sunt insolubili in apa si nu reactioneaza cu proteinele.
Orez. 1.11. Boabele complexe de aleuronă:
1 - pori în coajă; 2 - globoide; 3 - masa proteica amorfa; 4 - cristaloizi scufundați într-o masă proteică de amforă
Lipide de depozitare situate de obicei în hialoplasmă sub formă de picături și se găsesc în aproape toate celulele vegetale. Acesta este principalul tip de nutrienți de rezervă în majoritatea plantelor: semințele și fructele sunt cele mai bogate în ele. Grasimile (lipidele) sunt substanta cu cea mai mare rezerva calorica. Reactivul pentru substanțele asemănătoare grăsimilor este Sudan III, care le colorează portocaliu.
Carbohidrați sunt incluse în compoziția fiecărei celule sub formă de zaharuri solubile în apă (glucoză, fructoză, zaharoză) și polizaharide insolubile în apă (celuloză, amidon). În celulă, carbohidrații joacă rolul de sursă de energie pentru reacțiile metabolice. Zaharurile, atunci când sunt legate de alte substanțe biologice ale celulei, formează glicozide, iar polizaharidele cu proteine formează glicoproteine. Compoziția carbohidraților într-o celulă vegetală este mult mai diversă decât în celulele animale, datorită compoziției diverse a polizaharidelor din peretele celular și a zaharurilor din seva celulară a vacuolelor.
Principalul și cel mai comun carbohidrat de stocare este amidonul polizaharid. Amidonul asimilativ primar se formează în cloroplaste. Noaptea, când fotosinteza încetează, amidonul este hidrolizat în zaharuri și transportat în țesuturile de depozitare - tuberculi, bulbi, rizomi. Acolo, în tipuri speciale de leucoplaste - amiloplaste - unele dintre zaharuri se depun sub formă de boabe de amidon secundar. Boabele de amidon se caracterizează prin stratificare, care se explică prin conținut diferit de apă din cauza aprovizionării inegale de amidon în timpul zilei. Există mai multă apă în straturile întunecate decât în straturile luminoase. Un bob cu un centru de formare a amidonului în centrul amiloplastei se numește concentric simplu; dacă centrul este deplasat, se numește excentric simplu. Un bob cu mai mulți centri de formare a amidonului este complex. În boabele semicompuse, straturi noi sunt depuse în jurul mai multor centre de formare a amidonului, iar apoi straturi comune formează și acoperă centrii de formare a amidonului (vezi Fig. 1.12). Reactivul pentru amidon este o soluție de iod, care dă o culoare albastră.
Orez. 1.12. Boabele de amidon de cartofi (A):
1 - bob simplu; 2 - semicomplex; 3 - complex; grâu (B), ovăz (C)
Substante excretoare (produse metabolice secundare)
Incluziunile celulare includ, de asemenea, substanțe excretoare, de exemplu cristale de oxalat de calciu ( monocristale, rafide - cristale în formă de ac, druse - intercreșteri de cristale, nisip cristalin - o acumulare de multe cristale mici) (vezi Fig. 1.13). Mai rar, cristalele sunt compuse din carbonat de calciu sau silice ( cistoliti; vezi fig. 1.14). Cistoliții sunt depuși pe peretele celular, ieșind în celulă sub formă de ciorchini de struguri și sunt caracteristici, de exemplu, reprezentanților familiei de urzici și ai frunzelor de ficus.
Spre deosebire de animale, care excretă săruri în exces prin urină, plantele nu au organe excretoare dezvoltate. Prin urmare, se crede că cristalele de oxalat de calciu sunt produsul final al metabolismului protoplastelor, format ca un dispozitiv pentru îndepărtarea excesului de calciu din metabolism. De regulă, aceste cristale se acumulează în organele pe care planta le aruncă periodic (frunze, scoarță).

Orez. 1.13. Forme de cristale de oxalat de calciu în celule:
1, 2 - raphida (impatiens; 1 - vedere laterală, 2 - secțiune transversală); 3 - drusă (figur); 4 - nisip cristalin (cartofi); 5 - un singur cristal (vanilie)
Orez. 1.14. Cistolit (pe secțiunea transversală a unei frunze de ficus):
1 - pielea frunzei; 2 - cistolita
Uleiuri esentiale se acumuleaza in frunze (menta, lavanda, salvie), flori (macese), fructe (citrice) si seminte de plante (marar, anason). Uleiurile esențiale nu participă la metabolism, dar sunt utilizate pe scară largă în parfumerie (uleiuri de trandafir, iasomie), industria alimentară (uleiuri de anason, mărar), medicină (uleiuri de mentă, eucalipt). Rezervoarele pentru acumularea uleiurilor esențiale pot fi glande (mentă), recipiente lizigene (fructe citrice), fire de păr glandulare (mușcate).
Rășini- sunt compuși complecși formați în timpul vieții normale sau ca urmare a distrugerii țesuturilor. Ele sunt formate din celule epiteliale care căptușesc canalele de rășină ca un produs secundar al metabolismului, adesea cu uleiuri esențiale. Se pot acumula în seva celulară, citoplasmă sub formă de picături sau în recipiente. Sunt insolubile în apă, impermeabile la microorganisme și, datorită proprietăților lor antiseptice, cresc rezistența plantelor la boli. Rășinile sunt folosite în medicină, precum și la fabricarea vopselelor, lacurilor și uleiurilor lubrifiante. În industria modernă, acestea sunt înlocuite cu materiale sintetice.
Perete celular
Peretele celular rigid care înconjoară celula este alcătuit din microfibrile de celuloză încorporate într-o matrice care conține hemiceluloze și substanțe pectinice. Peretele celular oferă suport mecanic celulei, protejează protoplastul și menține forma celulei. În acest caz, peretele celular este capabil să se întindă. Fiind un produs al activității vitale a protoplastului, peretele poate crește doar în contact cu acesta. Apa și sărurile minerale se deplasează prin peretele celular, dar este complet sau parțial impermeabil la substanțele cu molecul mare. Când protoplastul moare, peretele poate continua să îndeplinească funcția de conducere a apei. Prezența unui perete celular, mai mult decât toate celelalte caracteristici, distinge celulele vegetale de celulele animale. Arhitectura peretelui celular este determinată în mare măsură de celuloză. Monomerul celulozei este glucoza. Mănunchiurile de molecule de celuloză formează micele, care se combină în fascicule mai mari - microfibrile. Reactivul pentru celuloză este clor-zinc-iod (Cl-Zn-I), care dă o culoare albastru-violet.
Cadrul celulozic al peretelui celular este umplut cu molecule ale matricei non-celulozice. Matricea contine polizaharide numite hemiceluloze; substanțe pectinice (pectină), foarte apropiate de hemiceluloze, și glicoproteine. Substanțele pectice, care se contopesc între celulele învecinate, formează o placă mediană, care este situată între membranele primare ale celulelor învecinate. Când placa de mijloc este dizolvată sau distrusă (care apare în pulpa fructelor coapte), are loc macerarea (din latinescul maceratio - înmuiere). Maceratul natural poate fi observat la multe fructe supracoapte (pepene verde, pepene galben, piersici). Macerația artificială (când țesutul este tratat cu alcali sau acid) este utilizată pentru prepararea diverselor preparate anatomice și histologice.
Peretele celular în procesul vieții poate suferi diverse modificări - lignificare, suberizare, mucilagii, cutinizare, mineralizare (vezi tabelul l.4).
Tabelul 1.4.
Informații conexe.