
Ce produce fitoplanctonul? Alge planctonice
Termen plancton(„planctonul” grecesc - rătăcire) a fost introdus pentru prima dată în știință de către Gezen în 1887 și, conform conceptului original, însemna o colecție de organisme care plutesc în apă. Ceva mai târziu, în compoziția planctonului au început să se distingă fitoplancton(plancton vegetal) și zooplancton(plancton animal). În consecință, fitoplanctonul este o colecție de plante mici, în principal microscopice, care plutesc liber (în coloana de apă), a căror mare parte sunt alge. În consecință, fiecare organism individual din compoziția fitoplanctonului este numit fitoplancter.
Ecologiștii consideră că fitoplanctonul în viața marilor corpuri de apă joacă același rol ca și plantele de pe uscat, adică produce materie organică primară, datorită căreia direct sau indirect (prin lanțul trofic) restul lumii vii există pe uscat. si in apa. Asta este adevărat. Cu toate acestea, trebuie amintit că fitoplanctonul, precum și comunitățile de plante terestre, includ ciuperci și bacterii, care, cu rare excepții, nu sunt capabile să creeze ele însele materie organică. Ei aparțin aceluiași grup ecologic de organisme heterotrofe care se hrănesc cu materie organică gata preparată, căruia îi aparține întreaga lume animală. Ciupercile și bacteriile participă la distrugerea materiei organice moarte, îndeplinind astfel, deși un rol foarte important în ciclul substanțelor, un rol fundamental diferit de plantele verzi. În ciuda acestui fapt, funcția principală a fitoplanctonului în general ar trebui totuși recunoscută ca fiind crearea de materie organică de către alge. Prin urmare, în continuare vom vorbi aici doar despre algele microscopice care fac parte din fitoplancton. Acest lucru este cu atât mai justificat cu cât compoziția ciupercilor din comunitatea fitoplanctonului este încă foarte puțin studiată, iar bacteriile planctonice (bacterioplanctonul) din ecologia corpurilor de apă sunt de obicei considerate separat.
Existenţa organismelor planctonice în suspensie în apă este asigurată de unele adaptări deosebite. La unele specii, se formează diferite tipuri de excrescențe și apendice ale corpului - spini, peri, procese asemănătoare cornului, membrane etc. (Fig. 27); la alte specii, în organism se acumulează substanțe cu greutatea specifică mai mică de unu, de exemplu, picături de grăsime, vacuole de gaz (la unele alge albastru-verzi, Fig. 28) etc. Masa celulei este, de asemenea, luminată. prin reducerea dimensiunii sale: dimensiunile celulelor la speciile planctonice, de regulă, sunt considerabil mai mici decât cele ale algelor de fund strâns înrudite. Cele mai mici organisme, de câțiva micrometri în dimensiune, formând așa-numitele nanoplancton.
, 
Compoziția și ecologia reprezentanților individuali ai fitoplanctonului algal din diferite corpuri de apă sunt extrem de diverse. Fitoplanctonul există în corpuri de apă de diferite naturi și dimensiuni - de la ocean până la o mică băltoacă. Este absent numai în rezervoare cu un regim brusc anormal, inclusiv în apele termale (la temperaturi ale apei peste +70, +80 °C), ape moarte (contaminate cu hidrogen sulfurat) și ape periglaciare curate care nu conțin substanțe nutritive minerale. De asemenea, nu există fitoplancton viu în lacurile din peșteri și la adâncimi mari de rezervoare, unde nu există suficientă energie solară pentru fotosinteză. Numărul total de specii de fitoplancton din toate apele marine și interioare ajunge la 3000.
În diferite corpuri de apă și chiar în același corp de apă, dar în diferite anotimpuri ale anului, numărul și raportul de specii ale grupurilor taxonomice individuale sunt foarte diferite. Să luăm în considerare principalele sale complexe în funcție de principalele categorii ecologice de corpuri de apă.
Fitoplanctonul marin constă în principal din diatomee și alge peridiniu. Folosirea metodelor de centrifugare și sedimentare a ajutat la descoperirea în plancton a unui număr semnificativ de specii de dimensiuni mici, necunoscute anterior. Dintre diatomeele din fitoplanctonul marin, reprezentanții clasei de diatomee centrice (Centrophyceae) sunt deosebit de numeroși, în special genurile Chaetoceros, Khizosolenia, Thalassiosira, Corethron, Planktoniella și unele altele (Fig. 29 , 1-6), complet absente din planctonul de apă dulce sau reprezentat în el doar de un număr mic de specii.
Compoziția formelor flagelate de alge pirofitice din fitoplanctonul marin este foarte diversă, mai ales din clasa peridinienilor (Fig. 29, 7-10). Acest grup este destul de divers în fitoplanctonul de apă dulce, dar are totuși mai puține specii decât în fitoplanctonul marin, iar unele genuri sunt reprezentate doar în mări: Dinophysis, Goniaulax și altele. De asemenea, foarte numeroase în fitoplanctonul marin sunt flagelate calcaroase - cocolitofore, reprezentate în apele dulci de doar câteva specii, și silicoflagelați, sau silicoflagelați, întâlniți exclusiv în planctonul marin (Tabelul 9).

Cea mai caracteristică trăsătură morfologică a reprezentanților fitoplanctonului marin este formarea diferitelor tipuri de excrescențe: peri și spini ascuțiți în diatomee, gulere, lobi și parașute în deridină. Formațiuni similare se găsesc și la speciile de apă dulce, dar acolo sunt mult mai puțin pronunțate. De exemplu, la speciile marine de Ceratium, procesele asemănătoare cornului sunt nu numai mult mai lungi decât la cele de apă dulce, dar la multe specii sunt și curbate. Se presupune că astfel de excrescențe contribuie la creșterea organismelor corespunzătoare. Conform altor idei, excrescențele precum țepii și formațiunile asemănătoare coarnelor s-au format ca un dispozitiv de protecție împotriva fitoplancterului consumat de crustacee și alți reprezentanți ai zooplanctonului.
Deși mediul marin este relativ omogen pe suprafețe mari, nu se observă o distribuție monotonă a fitoplanctonului. Eterogenitatea compoziției speciilor și diferențele de abundență sunt adesea pronunțate chiar și în zone relativ mici de apă de mare, dar sunt deosebit de pronunțate în distribuția geografică la scară largă. Aici se manifesta efectul ecologic al principalilor factori de mediu: salinitatea apei, temperatura, conditiile de iluminare si continutul de nutrienti.
Fitoplanctonul marin tropical se caracterizează prin cea mai mare diversitate de specii, în general cea mai scăzută productivitate (cu excepția zonelor de upwelling, care vor fi discutate mai jos) și cele mai pronunțate trăsături morfologice ale fitoplanctonului marin (diferitele tipuri de excrescențe menționate mai sus). Peridineii sunt extrem de diversi aici, printre care nu se numără doar specii individuale, ci și genuri întregi, distribuite exclusiv sau predominant în apele tropicale. Zona tropicala este biotopul optim (locul de existenta) iar pentru flagelate calcaroase - cocolitofore. Aici sunt cele mai diverse și în unele locuri se dezvoltă într-o astfel de masă încât scheletele lor calcaroase formează sedimente speciale de fund. Apele tropicale, în comparație cu apele reci din mările nordice și arctice, sunt mult mai sărace în diatomee. Verdele albastru, ca și în alte zone marine, sunt reprezentate de un număr foarte mic de specii, iar doar una dintre ele, aparținând genului Oscillatoria erythraea, se dezvoltă într-un asemenea număr în unele zone ale tropicelor încât provoacă „înflorirea” a apa.
Spre deosebire de tropice, în apele maritime polare și subpolare, fitoplanctonul este dominat de diatomee. Ei sunt cei care creează acea masă uriașă de produse vegetale lervine, pe baza căreia se formează acumulări puternice de zooplancton, care, la rândul său, servește drept hrană pentru cele mai mari turme de balene din Antarctica, hering și balene din apele polare ale Antarcticii. Arctic.
Peridinea în apele arctice sunt mult mai puțin reprezentate decât în mările de latitudini temperate și, mai ales, cele tropicale. Cocolitoforele sunt, de asemenea, rare aici, dar silicoflagelatele sunt diverse și pe alocuri numeroase. Algele marine albastre-verzi sunt absente, în timp ce unele tipuri de alge verzi se dezvoltă în cantități semnificative.
Nu mai puțin semnificative sunt diferențele în compoziția și productivitatea algelor în alte două biotopuri mari ale mărilor, delimitate pe direcția latitudinală - regiunile oceanice și neritice, mai ales dacă toate mările interioare sunt incluse în acestea din urmă. Caracteristicile speciale ale planctonului oceanic sunt enumerate mai sus. Deși sunt diferite în apele tropicale și subpolare, ele reflectă în general trăsăturile caracteristice ale fitoplanctonului marin. Planctonul oceanic, și numai acesta, este format exclusiv din specii care își încheie întregul ciclu de viață în coloana de apă - în zona pelagică a rezervorului, fără legătură cu solul. În planctonul neritic există deja semnificativ mai puține astfel de specii, iar în planctonul apelor continentale pot fi găsite doar ca excepție.
Zona neritică sau de raft este o zonă a mării care se extinde de la coastă până la capătul platformei continentale, care corespunde de obicei la o adâncime de aproximativ 200 m. În unele locuri este îngustă, în altele se extinde pe multe sute. și chiar mii de kilometri. Principalele caracteristici ecologice ale acestei zone sunt determinate de o legătură mai pronunțată cu malul și fundul. Aici există abateri semnificative de la condițiile oceanice în salinitatea apei (de obicei în jos); transparenta redusa datorita materiei minerale si organice in suspensie (adesea datorita productivitatii mai mari a planctonului); abateri ale conditiilor de temperatura; amestecare turbulentă mai pronunțată a apelor și, ceea ce este deosebit de important pentru planctonul vegetal, concentrația crescută de nutrienți.
Aceste caracteristici determină următoarele caracteristici în compoziția și productivitatea fitoplanctonului în zona neritică:
1) multe specii oceanice părăsesc această comunitate, altele sunt reprezentate în diferite grade prin forme modificate (soiuri);
2) apar multe specii marine specifice care nu se găsesc în planctonul oceanic;
3) se formează un complex de specii de apă salmastre care sunt complet absente în planctonul oceanic și în apele foarte desalinizate ale unor mări interioare, cu o salinitate a apei sub 10-12°/00 (°/00, ppm - o miime). de un număr, o zecime de procent) , speciile de apă dulce ating o diversitate semnificativă, care devin predominante atunci când apa este desalinizată la 2-3°/00;
4) apropierea fundului și țărmurilor contribuie la îmbogățirea fitoplanctonului neritic cu specii planctonice temporare (meroplanctonice).
Datorită diversității biotopilor, fitoplanctonul neritic în general este mult mai bogat în compoziția speciilor decât fitoplanctonul oceanic. Fitoplanctonul zonei neritice de latitudini temperate este dominat de diatomee și peridinieni, dar printre acestea se numără multe specii de apă salmastre, care se dezvoltă în mare parte în apele desalinizate ale mărilor interioare (Baltică, Neagră, Azov etc.). În ciclul de viață al multor specii de plancton neritic, este bine definită faza inferioară (etapa de repaus), ceea ce în latitudinile temperate determină o schimbare sezonieră (succesiune) mai clară a fitoplanctonului. În general, fitoplanctonul neritic este de câteva ori mai productiv decât fitoplanctonul oceanic.
Fitoplanctonul mărilor interioare desalinizate diferă semnificativ în compoziție și productivitate nu numai de planctonul oceanic, ci și de planctonul neritic tipic. Un exemplu este fitoplanctonul Mării Baltice. Salinitatea apei în stratul superior al părții centrale a Mării Baltice este de 7-8°/00, ceea ce este de aproximativ 4,5-5 ori mai mică decât salinitatea oceanului, dar de 20-40 de ori mai mare decât salinitatea apelor dulci. . În golfurile Riga, Finlanda și Botnia, salinitatea scade la 5-6°/00, în largul coastei - la 3-4°/00, și la gurile de râu și în unele golfuri estuare (Godul Neva, Laguna Curonian etc. ) apa este complet proaspătă.
Deși fitoplanctonul din Marea Baltică Centrală și chiar din partea deschisă a Golfului Riga, Finlanda și Botnia este dominat de un complex marin de specii, în sens strict nu poate fi numit marin decât prin origine. Speciile oceanice tipice sunt complet absente aici. Chiar și planctonul neritic marin este extrem de epuizat aici și este reprezentat doar de specii eurihaline - capabile să tolereze fluctuații mari de salinitate, deși preferând valori scăzute de salinitate. Acest complex fitoplancton baltic, de origine marină, dar salmastru ca ecologie, este dominat de specii de diatomee: Chaetoceros thalassiosira, Sceletonema, Actinocyclus. Peridineenii care se găsesc în mod regulat, dar nu ajung în număr mare, includ Goniaulax, Dinophysis baltica și câteva specii de silicoflagelate.
În fitoplanctonul Mării Baltice Centrale și în special în golfurile acesteia, un rol important îl joacă un complex de specii de origine de apă dulce, în principal albastru-verde: Anabaena, Aphanizomenon, Nodularia, Microcystis, care vara pe vreme însorită stabilă se dezvoltă într-un asemenea mod. masă care chiar și în partea centrală a mării formează o „înflorire” de apă (în principal datorită dezvoltării Aphanizomenon și Nodularia, iar în partea de sud a mării și Microcystis).
ÎN complex de apă dulce Algele verzi sunt si ele frecvente: Oocystis (pe toata marea), specii de Scenedesmus si Pediastrum, mai numeroase in golfuri.
Fitoplanctonul de apă dulce diferă de fitoplanctonul marin tipic într-o mare varietate de alge verzi și albastru-verzi. Deosebit de numeroase dintre cele verzi sunt speciile de volvox și protococi unicelulare și coloniale: specii de Chlamydomonas, Gonium, Volvox, Pediastrum, Scenedesmus, Oocystis, Sphaerocystis etc. (Fig. 30). Printre albastru-verzi se numără numeroase specii de anabena, microcystis, aphanizomenon, Gloeotrichia etc.

Diversitatea speciilor de diatomee aici este mai mică decât în mări (dacă nu luăm în considerare marea diversitate a speciilor planctonice temporare) (Fig. 31); În ceea ce privește productivitatea pe unitatea de suprafață a apei, rolul diatomeelor în apele dulci și în apele de mare este în medie comparabil.

Cel mai caracteristic gen de fitoplancton marin, Chaetoceros, este complet absent în lacuri și iazuri, iar Rhizosolenia, care este abundent în mări, este reprezentată în apele dulci de doar câteva specii.
În fitoplaktopul de apă dulce, peridinea este reprezentată într-o calitate și cantitate mult mai slabă. Des întâlnite printre ele sunt speciile de Ceratium și Peridinium, Fig. 64. În apele dulci, nu există flagelate silicioase și cocolitofore foarte rare, dar unele alte flagelate sunt reprezentate aici într-o varietate de moduri și adesea în număr mare. Acestea sunt în principal crisomonade - specii de Dinobryon, Mallomonas, Uroglena etc. (Fig. 68, 69), precum și euglena - Euglena, Trachelomonas și Phacus (Fig. 195, 201, 202); primul în principal în ape reci, iar cel din urmă în ape calde.
 ,
,  ,
,  ,
,  ,
,  ,
, 
Una dintre caracteristicile semnificative ale fitoplanctonului de apă dulce este abundența de alge planctonice temporare. Un număr de specii, care sunt considerate a fi de obicei planctonice, în iazuri și lacuri au o fază de fund sau perifitonă (atașare la orice obiect) în ciclul lor de viață. Astfel, diversitatea condițiilor de mediu din corpurile de apă interioară determină, de asemenea, o diversitate semnificativ mai mare a complexelor ecologice și a compoziției speciilor de plancton de apă dulce în comparație cu mările.
În lacurile mari și adânci, diferențele dintre fitoplanctonul de apă dulce și fitoplanctonul de mare sunt mai puțin pronunțate. În astfel de lacuri gigantice precum Baikal, Marile Lacuri, Ladoga, Onega, diatomeele predomină în fitoplancton aproape tot timpul anului. Aici, ca și în mări, ei creează principalele produse. Compoziția de specii a planctonului de lac diatomee este diferită de planctonul marin, dar ecologia lor are multe în comun. De exemplu, Melosira islandica, o specie de fitoplancton răspândită în lacurile Ladoga și Onega, precum și Melosira baicalensis din Lacul Baikal, în timpul fazei de repaus după focarul de primăvară, nu se scufundă în fund (sau doar parțial), așa cum este observate la alte specii de apă dulce în rezervoare mai mici, dar sunt reținute în coloana de apă, formând acumulări intersezonale caracteristice la o anumită adâncime. În lacurile mari, ca și în mări, există diferențe mari în productivitatea fitoplanctonului: în partea centrală a rezervorului, productivitatea este foarte scăzută, iar în largul coastei, în special în golfurile puțin adânci și împotriva gurilor de râu, crește brusc.
Fitoplanctonul celor mai mari două lacuri cu apă sărată din lume - Marea Caspică și Marea Aral - este și mai asemănător cu marea. Deși salinitatea apei în ele este semnificativ mai mică decât a apei de mare (în Marea Caspică 12-13°/00, în Marea Aral 11-120/00), compoziția fitoplanctonului de aici este dominată de algele de origine marina, în special în rândul diatomeelor. : specii de Chaetoceros, Rhizosolenia etc. Dintre flagelate, speciile tipice de apă salmastre sunt Exuviella și altele.În zonele desalinizate ale acestor lacuri predomină speciile de apă dulce, însă, la o salinitate a apei chiar de 3-5°/00, fitoplanctonul apelor salmastre de origine marina este inca foarte divers.
În forma sa cea mai tipică, fitoplanctonul de apă dulce, atât ca compoziție și ecologie, cât și ca proprietăți de producție, este reprezentat în lacurile de dimensiuni medii din zona temperată, de exemplu, în lacurile din bazinul baltic. Aici, în funcție de tipul de lac și de anotimpul anului, fitoplanctonul este dominat de diatomee, alge albastru-verzi sau verzi. Diatomeele tipice sunt Melosira, Asterionella, Tabellaria, Fragilaria, Cyclotella etc.; printre cele albastru-verzi se numără speciile de Microcystis, Anabaena, Aphanizomenon și Gloeotrichia. Principalii reprezentanți ai algelor verzi din planctonul lacului sunt cele protococice enumerate mai sus, iar în apele cu apă foarte moale, sub influența mlaștinilor, desmididele sunt numeroase: specii de Cosmarium, Staurastrum, Closterium, Euastrum etc. În lacurile de mică adâncime și iazuri, algele verzi sunt adesea dominate de Volvox: Volvox, Chlamydomonas, Pandorina, Eudorina. În fitoplanctonul lacurilor din tundra și taiga de nord, chrysomonas sunt foarte diverse: specii de Dinobryon, Synura, Uroglenopsis, Mallomonas. Grupa peridinei, cea mai caracteristică fitoplanctonului marin, este reprezentată în apele dulci de pretutindeni (în toate corpurile de apă), dar de un număr relativ mic de specii, care peste tot, cu rare excepții, ajung la un număr redus. În cele mai mici corpuri de apă - în lacuri și iazuri mici - euglene sunt foarte diverse și adesea numeroase, în special specii de Trachelomonas, iar în rezervoarele calde ale tropicelor și subtropicelor se găsesc și euglene, lepocynclis, Phacus etc.
În fiecare rezervor individual, în funcție de caracteristicile fizico-chimice ale regimului și anotimpul anului, predomină una sau alta dintre grupele de alge enumerate, iar în perioadele de dezvoltare foarte intensă, de multe ori domină doar o singură specie.
În rezervoarele temporare mici - bălți, gropi săpate - speciile mici de volvox din genul Chlamydomonas sunt destul de comune, a căror dezvoltare în masă deseori înverzește apa.
În literatură, fitoplanctonul fluvial este adesea clasificat ca o categorie specială de plancton de apă dulce. În râurile mari cu debite foarte lente, desigur, algele au timp să se înmulțească într-o zonă limitată a râului în condiții relativ uniforme. În consecință, aici se poate forma o compoziție de fitoplancton care este oarecum unică pentru aceste condiții. Cu toate acestea, chiar și în acest caz, „materialul” inițial pentru o anumită comunitate fluvială este organisme transportate de curent dintr-o secțiune din amonte a râului sau din afluenții laterali. Cel mai adesea, compoziția fitoplanctonului dintr-un râu se formează ca un amestec de fitoplancton al afluenților, transformat într-un grad sau altul sub influența condițiilor râului.
Rolul transformator al condițiilor râului în formarea fitoplanctonului său este clar demonstrat atunci când un râu mare de câmpie curge printr-un oraș sau pe lângă o centrală mare care poluează apa cu ape uzate menajere și industriale. În acest caz, compoziția fitoplanctonului din râul de deasupra orașului caracterizează apa curată, dar în interiorul orașului și imediat dincolo de periferia acestuia, sub influența poluării organice, fitoplanctonul este foarte epuizat și predomină așa-numitele specii saprobice - indicatori de saprobic. , adică ape poluate. Totuși, dedesubt, parțial datorită sedimentării materiei organice în suspensie, parțial datorită dezintegrarii acestora ca urmare a proceselor microbiologice, apa redevine limpede, iar fitoplanctonul capătă aproximativ același aspect ca și deasupra orașului.
Compoziția și distribuția fitoplanctonului în rezervoare individuale și modificările sale într-un singur rezervor sunt influențate de un complex mare de factori. De importanță primordială printre factorii fizici sunt regimul de lumină, temperatura apei și pentru rezervoarele adânci - stabilitatea verticală a maselor de apă. Dintre factorii chimici, importanța principală este salinitatea apei și conținutul de nutrienți din aceasta, în primul rând săruri de fosfor, azot, iar pentru unele specii și fier și siliciu. Să ne uităm la câțiva dintre acești factori.
Influența iluminării ca factor de mediu se manifestă clar în distribuția verticală și sezonieră a fitoplanctonului. În mări și lacuri, fitoplanctonul există doar în stratul superior al apei. Limita sa inferioară în mare, ape mai transparente se află la adâncimea de 40-70 m și doar în câteva locuri ajunge la 100-120 m (Marea Mediterană, apele tropicale ale Oceanului Mondial). În apele lacurilor, care sunt mult mai puțin transparente, fitoplanctonul există de obicei în straturile superioare, la adâncimea de 10-15 m, iar în apele cu transparență foarte scăzută se găsește la adâncimea de 2-3 m. Numai în înaltă- munte și unele lacuri mari (de exemplu, Baikal) cu În apa limpede, fitoplanctonul este distribuit la o adâncime de 20-30 m. În acest caz, transparența apei afectează algele nu direct, ci indirect, deoarece determină intensitatea pătrunderii radiația solară în coloana de apă, fără de care fotosinteza este imposibilă. Această fântână confirmă cursul sezonier al dezvoltării fitoplanctonului în corpurile de apă de latitudini temperate și înalte, care îngheață iarna. În timpul iernii, când rezervorul este acoperit cu gheață, adesea cu un strat de zăpadă, în ciuda celei mai mari transparențe a apei a anului, fitoplanctonul este aproape absent - se găsesc doar foarte rare celule inactive fiziologic ale unor specii, iar la unele alge - spori. sau celule în stadiu de repaus.
Având în vedere dependența generală mare a fitoplanctonului de iluminare, valorile optime ale acestuia din urmă pentru specii individuale variază într-un interval destul de larg. Algele verzi și majoritatea speciilor de alge albastre-verzi, care se dezvoltă în număr semnificativ în sezonul estival, sunt deosebit de solicitante față de acest factor. Unele specii de albastru-verzi se dezvoltă în masă doar la suprafața apei: Oscillatoria - în mările tropicale, multe specii de Microcystis, Anabaena etc. - în apele interioare puțin adânci.
Diatomeele sunt mai puțin solicitante în condițiile de iluminare. Majoritatea lor evită stratul de suprafață puternic luminat de apă și se dezvoltă mai intens doar la o adâncime de 2-3 m în apele puțin transparente ale lacurilor și la o adâncime de 10-15 m în apele limpezi ale mărilor.
Temperatura apei este cel mai important factor în distribuția geografică generală a fitoplanctonului și a ciclurilor sale sezoniere, dar în multe cazuri acest factor acționează nu direct, ci indirect. Multe alge sunt capabile să tolereze o gamă largă de fluctuații de temperatură (specii euritermale) și se găsesc în plancton de diferite latitudini geografice și în diferite anotimpuri ale anului. Cu toate acestea, zona optimă de temperatură, în cadrul căreia se observă cea mai mare productivitate, pentru fiecare specie este de obicei limitată de mici abateri de temperatură. De exemplu, diatomea Melosira islandica, răspândită în planctonul lacului din zona temperată și subarctică, este de obicei prezentă în plancton (de exemplu, în lacurile Onega și Ladoga, în Neva) la temperaturi de la +1 la + 13 ° C și reproducerea sa maximă se observă la temperaturi de la +6 la +8 °C.
Temperatura optimă pentru diferite specii nu coincide, ceea ce determină modificarea compoziției speciilor de-a lungul anotimpurilor, așa-numita succesiune sezonieră a speciilor. Schema generală a ciclului anual al fitoplanctonului în lacurile de latitudini temperate este următoarea. Iarna, sub gheață (mai ales când gheața este acoperită cu zăpadă), fitoplanctonul este aproape absent din cauza lipsei radiației solare. Ciclul de vegetație al fitoplanctonului ca comunitate începe în martie - aprilie, când radiația solară este suficientă pentru fotosinteza algelor chiar și sub gheață. În acest moment, flagelate mici - Cryptomonas, Chromulina, Chrysococcus - sunt destul de numeroase - iar numărul speciilor de diatomee de apă rece - Melosira, Diatoma etc. - începe să crească.
În a doua fază a primăverii - din momentul în care gheața se sparge pe lac și până la stabilirea stratificării temperaturii, ceea ce se întâmplă de obicei atunci când stratul superior de apă se încălzește până la +10, +12 °C, o dezvoltare rapidă a frigului - se observă complexul de diatomee de apă. În prima fază a sezonului de vară, la temperaturi ale apei de la +10 la + 15 °C, complexul de apă rece de diatomee încetează să crească.În acest moment, diatomeele sunt încă numeroase în plancton, dar alte specii sunt moderat calde. apă: Asterionella, Tabellaria . În același timp, crește productivitatea algelor verzi și albastre-verzi, precum și a crisomonadelor, unele specii atingând o dezvoltare semnificativă deja în a doua fază a primăverii. În a doua fază a verii, la temperaturi ale apei peste + 15 °C, se observă productivitatea maximă a algelor albastre-verzi și verzi. În funcție de tipul trofic și limnologic al rezervorului, în acest moment poate exista o „înflorire” a apei cauzată de specii de alge albastru-verzi (Anabaena, Aphanizomenon, Microcystis, Gloeotrichia, Oscillatoria) și verzi (Scenedesmus, Pediastrum, Oocystis). ).
Vara, diatomeele, de regulă, ocupă o poziție subordonată și sunt reprezentate de specii de apă caldă: Fragilaria și Melosira granulata. Toamna, cu o scădere a temperaturii apei la +10, +12 °C și mai jos, se observă din nou o creștere a productivității speciilor de diatomee de apă rece. Cu toate acestea, spre deosebire de sezonul de primăvară, algele albastre-verzi joacă un rol semnificativ mai mare în acest moment.
În apele mării de latitudini temperate, faza de primăvară în fitoplancton este marcată și de un focar de diatomee; vara - o creștere a diversității speciilor și a abundenței peridinei cu o scădere a productivității fitoplanctonului în general.
Printre factorii chimici care influențează distribuția fitoplanctonului, pe primul loc trebuie pusă compoziția de sare a apei. În același timp, concentrația totală de săruri este un factor important în distribuția calitativă (speciilor) între tipurile de rezervoare, iar concentrația de săruri nutritive, în principal săruri de azot și fosfor, este o distribuție cantitativă, adică productivitate.
Concentrația totală de săruri ale apelor naturale normale (în sens ecologic) variază într-un interval foarte larg: de la aproximativ 5-10 la 36.000-38.000 mg/l (de la 0,005-0,01 la 36-38°/0O). În acest interval de salinitate, se disting două clase principale de corpuri de apă: marine cu o salinitate de 36-38°/00, adică 36.000-38.000 mg/l, și proaspătă cu o salinitate de la 5-10 la 400-500 și chiar în sus. la 1000 mg/l. Apele salmastre ocupă o poziție intermediară în ceea ce privește concentrația de sare. Aceste clase de ape, așa cum se arată mai sus, corespund și principalelor grupuri de fitoplancton în ceea ce privește compoziția speciilor.
Semnificația ecologică a concentrației de nutrienți se manifestă în distribuția cantitativă a fitoplanctonului în ansamblu și a speciilor sale constitutive.
Productivitatea, sau „randamentul” algelor microscopice fitoplancton, ca și randamentul vegetației mari, în alte condiții normale, depinde într-o foarte mare măsură de concentrația de nutrienți din mediu. Dintre nutrienții minerali pentru alge, ca și pentru vegetația terestră, sunt necesare în primul rând sărurile de azot și fosfor. Concentrația medie a acestor substanțe în majoritatea corpurilor naturale de apă este foarte mică și, prin urmare, productivitatea ridicată a fitoplanctonului, ca fenomen stabil, este posibilă numai dacă substanțele minerale intră constant în stratul superior al apei - zona fotosintezei.
Adevărat, unele alge albastre-verzi sunt încă capabile să absoarbă azotul elementar din aerul dizolvat în apă, dar există puține astfel de specii și rolul lor în îmbogățirea cu azot este semnificativ doar pentru corpurile de apă foarte mici, în special în câmpurile de orez.
Rezervoarele interioare sunt fertilizate cu azot și fosfor de pe țărm, datorită aprovizionării cu nutrienți de către apa râului din zona de drenaj a întregului sistem fluvial. Prin urmare, există o dependență clară a productivității lacurilor și mărilor interioare de mică adâncime de fertilitatea solului și de alți factori care operează în zona de drenaj a bazinelor lor (sistemele fluviale). Fitoplanctonul cel mai puțin productiv este în lacurile periglaciare, precum și în rezervoarele situate pe roci cristaline și în zonele cu un număr mare de mlaștini din zona de drenaj. Exemple ale acestora din urmă sunt lacurile din Karelia de Nord, Peninsula Kola, Finlanda de Nord, Suedia și Norvegia. Dimpotrivă, rezervoarele situate în soluri foarte fertile se caracterizează printr-un nivel ridicat de productivitate a fitoplanctopurilor și a altor comunități (Marea Azov, rezervoarele Volga de Jos, rezervorul Tsimlyansk).
Productivitatea fitoplanctonului depinde și de dinamica apei și de regimul dinamic al apei. Influența poate fi directă și indirectă, ceea ce, totuși, nu este întotdeauna ușor de distins. Amestecarea turbulentă, dacă nu este prea intensă, în alte condiții favorabile, contribuie direct la creșterea productivității diatomeelor, deoarece multe specii din această diviziune, având o înveliș relativ grea de siliciu, se scufundă până la fund în apă calmă. Prin urmare, o serie de specii abundente de apă dulce, în special din genul Melosira, se dezvoltă intens în planctonul lacurilor de latitudini temperate numai primăvara și toamna, în perioadele de amestec vertical activ al apei. Când o astfel de amestecare încetează, ceea ce are loc atunci când stratul superior se încălzește până la +10, +12 °C și se formează stratificarea temperaturii coloanei de apă în multe lacuri, aceste specii ies din plancton.
Alte alge, în principal alge albastre-verzi, dimpotrivă, nu pot tolera nici măcar amestecarea relativ slabă a apei turbulente. Spre deosebire de diatomee, multe specii albastru-verde se dezvoltă cel mai intens în apă extrem de calmă. Motivele sensibilității lor ridicate la dinamica apei nu sunt pe deplin stabilite.
Cu toate acestea, în cazurile în care amestecarea verticală a apelor se extinde la adâncimi mari, suprimă dezvoltarea chiar și a diatomeelor relativ tolerante la umbră. Acest lucru se datorează faptului că în timpul amestecării profunde, algele sunt transportate periodic de curenții de apă în afara zonei iluminate - zona de fotosinteză.
Influența indirectă a factorului dinamic asupra productivității fitoplanctonului este aceea că, prin amestecarea verticală a apei, nutrienții se ridică din straturile inferioare ale apei, unde nu pot fi folosiți de alge din cauza lipsei de lumină. Aici se manifestă interacțiunea mai multor factori de mediu - regimuri luminoase și dinamice și aprovizionarea cu nutrienți. Această relație este tipică pentru procesele naturale.
Deja la începutul acestui secol, hidrobiologii au descoperit importanța deosebită a fitoplanctonului în viața corpurilor de apă ca principală, iar în vastele întinderi oceanice, singurul producător de materie organică primară, pe baza căruia restul diversității. a vieţii acvatice este creată. Acest lucru a determinat un interes sporit pentru studierea nu numai a compoziției calitative a fitoplanctonului, ci și a distribuției sale cantitative, precum și a factorilor care reglează această distribuție.
O metodă elementară de cuantificare a fitoplanctonului, care a fost metoda principală de câteva decenii și nu a fost încă complet abandonată, este metoda de strecurare a acestuia din apă folosind grile de plancton. Într-o probă concentrată în acest mod, se calculează numărul de celule și colonii pe specii și se determină numărul total al acestora pe unitatea de suprafață a rezervorului. Această metodă simplă și accesibilă are însă un dezavantaj semnificativ - nu ia în considerare pe deplin nici măcar algele relativ mari, iar cele mai mici (nannoplancton), care predomină semnificativ în multe rezervoare, nu sunt captate de plasele de plancton.
În prezent, probele de fitoplancton sunt prelevate în principal cu un batometru sau planktobatometru, ceea ce face posibilă „decuparea” unui monolit de apă de la o anumită adâncime. Proba se concentrează prin sedimentare în cilindri sau prin filtrare prin microfiltre: ambele asigură că sunt luate în considerare algele de toate dimensiunile.
Când au fost determinate diferențe uriașe în dimensiunile algelor care alcătuiesc fitoplanctonul (de la câțiva la 1000 de microni sau mai mult), a devenit clar că valorile abundenței nu pot fi folosite pentru o evaluare comparativă a productivității fitoplanctonului în corpurile de apă. Un indicator mai realist în acest scop este biomasa totală de fitoplancton pe unitatea de suprafață a rezervorului. Cu toate acestea, mai târziu, această metodă a fost respinsă din două motive principale: în primul rând, calculele biomasei celulelor care au configurații diferite în diferite specii necesită foarte multă muncă; în al doilea rând, contribuția algelor mici, dar care se reproduc rapid, la producția totală a comunității pe unitatea de timp poate fi semnificativ mai mare decât cea a algelor mari, dar care se reproduc încet.
Adevăratul indicator al productivității fitoplanctonului este rata de formare a materiei pe unitatea de timp. Pentru a determina această valoare, se utilizează o metodă fiziologică. În timpul procesului de fotosinteză, care are loc numai în lumină, dioxidul de carbon este absorbit și oxigenul este eliberat. Odată cu fotosinteza are loc și respirația algălui. Ultimul proces, asociat cu absorbția oxigenului și eliberarea de dioxid de carbon, predomină în întuneric, când fotosinteza se oprește. Metoda de evaluare a productivității fitoplanctonului se bazează pe o comparație cantitativă a rezultatelor fotosintezei (procesul de producție) și respirației (procesul de distrugere) a comunității pe baza echilibrului de oxigen din rezervor. În acest scop, probele de apă sunt folosite în sticle deschise și întunecate, expuse într-un rezervor, de obicei pentru o zi la diferite adâncimi.
Pentru a crește sensibilitatea metodei oxigenului, care este nepotrivită pentru apele neproductive, au început să folosească o versiune izotopică (radiocarbon) a acesteia. Cu toate acestea, ulterior au fost relevate deficiențele metodei oxigenului în ansamblu, iar în prezent este utilizată pe scară largă metoda clorofilei, bazată pe determinarea conținutului de clorofilă într-o probă cantitativă de fitoplancton.
În prezent, nivelul productivității fitoplanctonului în multe corpuri de apă interioară este determinat nu atât de condițiile naturale, cât de condițiile socio-economice, adică de densitatea populației și natura activității economice din zona de captare a lacului de acumulare. Această categorie de factori, numiți antropici în ecologie, adică proveniți din activitatea umană, duce la epuizarea fitoplanctonului în unele corpuri de apă, iar în altele, dimpotrivă, la o creștere semnificativă a productivității acestuia. Prima apare ca urmare a deversarii intr-un rezervor a substantelor toxice continute in apele uzate industriale, iar a doua apare atunci cand rezervorul este imbogatit cu substante nutritive (in special compusi fosforici) in forma minerala sau organica, continuti in concentratii mari in apele care curg din zone agricole și orașe și sate mici (ape uzate menajere). Nutrienții se găsesc și în apele uzate din multe procese industriale.
Al doilea tip de influență antropică - îmbogățirea unui rezervor cu substanțe nutritive - crește productivitatea nu numai a fitoplanctonului, ci și a altor comunități acvatice, inclusiv a peștilor, și ar trebui considerat ca un proces favorabil din punct de vedere economic. Cu toate acestea, în multe cazuri, îmbogățirea antropică spontană a corpurilor de apă cu nutrienți primari are loc la o asemenea scară încât corpul de apă ca sistem ecologic devine supraîncărcat cu nutrienți. Consecința acestui lucru este dezvoltarea excesiv de rapidă a fitoplanctonului („înflorirea” apei), a cărui descompunere eliberează hidrogen sulfurat sau alte substanțe toxice. Acest lucru duce la moartea populației animale din rezervor și face ca apa să nu fie aptă de băut.
Există și cazuri frecvente de eliberare intravitală de substanțe toxice de către alge. În corpurile de apă dulce, acest lucru se observă cel mai adesea cu dezvoltarea masivă a algelor albastre-verzi, în special speciile din genul Microcystis. În apele mării, otrăvirea apei este adesea cauzată de dezvoltarea masivă a flagelaților mici. În astfel de cazuri, apa devine uneori roșie, de unde și numele acestui fenomen - „marea roșie”.
O scădere a calității apei ca urmare a supraîncărcării antropice a unui rezervor cu substanțe nutritive, care determină dezvoltarea excesivă a fitoplanctonului, este de obicei numită fenomenul de eutrofizare antropică a unui rezervor. Aceasta este una dintre manifestările triste ale poluării umane a mediului. Amploarea acestui proces poate fi judecată după faptul că poluarea se dezvoltă intens în corpuri de apă dulce atât de uriașe precum Lacul Erie și chiar în unele mări.
Fertilitatea naturală a apelor de suprafață a mării este determinată de diverși factori. Reaprovizionarea cu nutrienți în mările interioare de mică adâncime, de exemplu în Marea Baltică și Azov, are loc în principal datorită alimentării lor de către apele râurilor.
Apele de suprafață ale oceanelor sunt îmbogățite cu substanțe nutritive în zonele unde apele adânci ajung la suprafață. Acest fenomen este inclus în literatură sub denumirea de upwelling. Upwelling-ul este foarte intens în largul coastei peruane. Pe baza producției mari de fitoplancton, producția de nevertebrate este extrem de mare aici, iar din această cauză crește numărul de pești. O țară mică, Peru în anii 60 a ocupat primul loc în lume în ceea ce privește capturile de pește.
Productivitatea puternică a fitoplanctonului în apele reci ale mărilor arctice și mai ales în apele Antarcticii este determinată și de creșterea apelor adânci îmbogățite cu substanțe nutritive. Un fenomen similar este observat în alte zone ale oceanului. Fenomenul opus, adică epuizarea apelor de suprafață în nutrienți, care inhibă dezvoltarea fitoplanctonului, se observă în zonele cu izolarea stabilă a apelor de suprafață de apele adânci.
Acestea sunt principalele caracteristici ale fitoplanctonului tipic.
Printre comunitățile de plante și animale mici care locuiesc în coloana de apă, există un complex de organisme care trăiesc doar la suprafața apei - în zona peliculei de suprafață. În 1917, Nauman a dat acestei comunități, nu atât de semnificativă în ceea ce privește compoziția speciilor, ci unei comunități foarte unice, un nume special - Neuston(greacă „nein” - a înota), deși, evident, este doar o parte integrantă a planctonului.
Viața organismelor neuston este asociată cu pelicula de suprafață a apei, iar unele dintre ele sunt situate deasupra peliculei (epineuston), altele - sub film (hyponeuston). Pe lângă algele și bacteriile microscopice, aici trăiesc și animale mici - nevertebrate și chiar larvele unor pești.
Concentrații mari de organisme neuston au fost găsite inițial în corpuri mici de apă - în iazuri, gropi săpate, în golfuri mici ale lacurilor - pe vreme calmă, cu o suprafață de apă calmă. Mai târziu, o varietate de organisme neuston, mai ales animale mici, au fost găsite în corpuri mari de apă, inclusiv în mări.
Compoziția algelor neuston de apă dulce include specii din diferite grupuri sistematice. Aici s-au găsit o serie de reprezentanți ai algelor aurii - Chromulina, Kremastochrysis; din euglena - euglena (Euglena), trachelomonas (Tgachelomonas), precum și unele verzi - chlamydomonas (Chlamydomonas), kremastochloris (Kremastochloris) - și mici protococi, anumite specii de galben-verzui și diatomee.
Unele specii de alge neuston au adaptări caracteristice pentru a exista la suprafața apei. De exemplu, speciile de Nautococcus au parașute de mucus care le țin de pelicula de suprafață. În Cremastochrysis (Fig. 32, 1), pentru aceasta se folosește o parașută solzoasă; la o specie de alge verzi, o astfel de parașută microscopică iese deasupra filmului de tensiune superficială sub forma unui capac în formă de con (Fig. 32, 2).

Avantajele existenței organismelor neuston la limita mediului de apă și aer sunt neclare, totuși, în unele cazuri se dezvoltă într-un asemenea număr încât acoperă apa cu o peliculă continuă. Adesea, algele planctonice (în special algele albastre-verzi) în timpul perioadei de dezvoltare în masă plutesc până la suprafața apei, formând acumulări uriașe. Au fost găsite și concentrații puternic crescute de bacterii acvatice. În comunitatea neuston, animalele microscopice sunt, de asemenea, destul de diverse, care, chiar și în mări, în condițiile unei suprafețe aproape constant turbulente, formează uneori acumulări semnificative la marginea inferioară a suprafeței apei.
Enciclopedia geologică Wikipedia - Acest articol trebuie rescris complet. S-ar putea să fie explicații pe pagina de discuții... Wikipedia
Clasificare științifică Regatul: Chromists ... Wikipedia
Partea vegetală a planctonului este distribuită în stratul de apă (în medie 200 m în Oceanul Mondial) care primește energie solară (zona eufotică). Fitoplanctonul este principalul producător primar de materie organică din corpurile de apă, datorită... ... Dicționar ecologic
fitoplancton- O parte a planctonului reprezentată de plante. [GOST 30813 2002] fitoplancton Alge unicelulare care trăiesc în stratul iluminat superior al apei. [Dicționar de termeni și concepte geologice. Universitatea de Stat din Tomsk] Subiecte: alimentare cu apă și... Ghidul tehnic al traducătorului
FITOPLANCTON- (din fito... și plancton) o colecție de plante microscopice (în principal alge) care trăiesc în grosimea apelor mari și dulci și se mișcă pasiv sub influența curenților de apă. O sursă de materie organică într-un corp de hrană pentru alții... ... Dicţionar enciclopedic mare
FITOPLANCTON- PHYTOPLANKTON, o colecție de mici plante oceanice care derivă odată cu curentul, spre deosebire de ZOOPLANKTON, o colecție de mici organisme animale care derivă odată cu curentul. Majoritatea fitoplanctonului au dimensiuni microscopice, de exemplu... Dicționar enciclopedic științific și tehnic
fitoplancton- substantiv, număr de sinonime: 1 microfitoplancton (1) Dicționar de sinonime ASIS. V.N. Trishin. 2013… Dicţionar de sinonime
FITOPLANCTON- o colecție de alge care trăiesc în stratul de apă iluminat superior. F. formează alge unicelulare descompuse. apartenența sistematică sunt aurii, peridinia, diatomeele, albastru-verde, heteroflagelate, euglenaceae etc., având un număr de... ... Enciclopedie geologică
Fitoplancton- un set de plante unicelulare care trăiesc în stratul fotic al oceanului. Este principala sursă de noi formări de materie organică în ocean. Îngreunează detectarea submarinelor. EdwART. Dicţionar naval explicativ, 2010 ... Dicţionar marin
fitoplancton- Un set de organisme vegetale care alcătuiesc planctonul (diatomee, alge verzi și albastre-verzi)... Dicţionar de Geografie
FITOPLANCTON- organisme vegetale care plutesc liber (alge) care locuiesc în straturile de suprafață ale apei. Dezvoltarea masivă a fosforului în iazuri conferă apei o anumită culoare. F. este o sursă de producție primară (materie organică) și o sursă de oxigen... ... Piscicultură de iaz
Cărți
- Fitoplanctonul rezervoarelor Volga de Jos și cursurile inferioare ale râului, Trifonova I. (ed.). Nu există un sistem unificat general acceptat pentru analiza biologică a calității apei. O scurtă analiză a situației ecologice din bazinul hidrografic. Volga și alte râuri arată nevoia de a efectua... Cumpărați pentru 151 de ruble
- Fitoplanctonul Volga de Jos. Rezervor și cursuri inferioare ale râului. Cartea prezintă caracteristicile limnologice ale rezervoarelor Volga de Jos - Kuibyshev, Saratov și Volgograd, precum și caracteristicile fizice și geografice ale regiunii în ansamblu. Dat...
Ciclul de producere a fitoplanctonului
Principalii factori care determină producția primară de fitoplancton sunt grosimea stratului fotic, aprovizionarea cu fitoplancton cu nutrienți și durata ciclului de producție.
În general, acești parametri depind de condițiile climatice și hidrologice specifice fiecărei regiuni și se manifestă în momentul începerii dezvoltării primăverii, care este strâns legat de regimul de urme și de sfârșitul ciclului de producție; în plus, acești parametri sunt determinați de aportul de nutrienți formați în stratul fotic la începutul dezvoltării primăverii și determinați de scurgerea râului și de adâncimea amestecării convective de iarnă, difuzia turbulentă verticală de vară, cantitatea de energie solară (în funcție de rândul ei). privind perioada anului și factorii climatici), precum și caracteristicile structurale și funcționale ale fitoplanctonului.
Fitoplanctonul Mării Albe este planctonul sărăcit calitativ al Mării Barents, cu un amestec mare de forme epifitice „litorale” și de apă dulce (Kokin, Koltsova, 1972). În Marea Albă, comparativ cu partea de sud a Mării Barents, numărul speciilor este dominat de diatomee iubitoare de frig, iar diversitatea speciilor de peridini este redusă. Tivul conținutului de alge din compoziția fitoplanctonului (63%) a Mării Albe este aproape de mările epicontinentale ale arctice înalte precum Kara (67%) și Laptev (64%), ceea ce indică încă o dată natura sa arctică. Formarea compoziției calitative și cantitative a fitoplanctonului în diferite anotimpuri biologice are loc sub influența factorilor de mediu atât biotici, cât și abiotici.
Modificările parametrilor structurali și funcționali ai comunității în timpul ciclului de producție pot fi urmărite prin exemplul analizei anotimpurilor biologice din Golful Kandalaksha în 1970 (Fedorov și Bobrov, 1977, Bobrov, 1982).
În total, în perioada de observație de la 15 aprilie până la 26 octombrie 1970, au fost instalate 51 de stații (de două ori pe săptămână). Produsele au fost determinate folosind metoda radiocarbonului baloanelor la orizonturi de 0,2,5, 5, 10, 25, 50, 75 m și la fund. Radiația solară totală a fost măsurată cu un piranometru Janyshevsky într-o modificare subacvatică. Concomitent cu experimentele de determinare a producției primare din orizonturile corespunzătoare, s-au efectuat colecții batometrice cantitative de fitoplancton și studiul unor parametri hidrologici și hidrochimici. La calcularea eficienței procesului fotosintetic, aceste măsurători ale producției primare au fost exprimate în unități energetice, pe baza următorului calcul: se consumă 112 calorii de energie luminoasă pentru a fixa 1 mol de CO2, iar această valoare corespunde la 9,36 cal/mgC.
Trecerea de la numărul de indivizi fitoplancton la caracteristicile de greutate s-a făcut după formula:
Formula a fost obținută ținând cont de faptul că biomasa uscată de alge conține 35% cenușă (Fedorov și colab., 1974).
Valorile cantităților zilnice de radiații în timpul sezonului de vegetație din 1970 au variat între 20,6 și 2300 cal/(cm") zi. Creșterea radiației solare care a pătruns în BOAy în primele zece zile ale lunii mai până la 100,6-181,4 cal/( cm" zi) coincide cu ruperea stratului de gheață. O ușoară scădere a valorilor energiei solare totale a fost cauzată de condițiile meteorologice.Îmbunătățirea condițiilor meteorologice și creșterea duratei orelor de lumină au dus la coincidența practică a maximului luminos (2300 cal/(cm. zi)) cu ziua calendaristică a solstițiului de vară.Scăderea ulterioară a cantităților zilnice de radiație solară are un caracter destul de neted și este cauzată de o scădere a duratei LEHI HEM CBPTOBOTO IH H H BECOTE CONHL kl Până la sfârșitul sezonului de vegetație, cantitatea de energie solară scade la 20 cal/(cm. zi).
Astfel, pentru întreaga perioadă de observare, care acoperă 183 de zile, valoarea energetică a fost exprimată ca valoare totală de 22,8 kcal/cm Energia necontabilizată a radiației solare penetrante în perioada de toamnă a „izvorului subglaciar” a fost obținută prin extrapolare și a fost însumată până la 22 kcal/cm.” În total, 25 kcal/cm au pătruns în rezervor în timpul sezonului." Această din urmă valoare poate fi considerată o sumă anuală, deoarece în lunile de iarnă cantitatea de energie de radiație solară care pătrunde sub stratul de gheață este nesemnificativă. Studiul distribuției. a energiei solare în coloana de apă în perioada de vegetație a făcut posibilă calcularea coeficientului de absorbție a luminii cu apă k=0,37-002.
Temperatura din orizontul de suprafață în timpul sezonului de vegetație a variat între -1,54 și 15,65 °C. Fluctuațiile de temperatură s-au atenuat cu adâncimea. Perioada de primăvară după spargerea gheții este caracterizată de o încălcare a homotermiei și de stabilirea unei stratificări clar definite. Zona de salt de temperatură, cu o creștere a energiei solare penetrante și încălzirea corespunzătoare a VM, s-a mutat treptat de la suprafață la straturile mai profunde. În perioada de maximă energie solară, termoclinul a fost situat la o adâncime de 25 m și a rămas la acest orizont până la sfârșitul lunii septembrie. În octombrie, saltul de temperatură a fost localizat mai adânc și s-a dezvăluit o tendință spre homeotermie.
În tabel Tabelul 5.1 prezintă câteva caracteristici abiotice ale Mării Albe, obținute pe baza analizei LHHaMHK.H sezoniere ale acestora.
Analiza dezvoltării vegetative a planctonului și a producției sale HEIM Xapa KTepH cTH K face posibilă identificarea și stabilirea duratei principalelor perioade de vegetație, numite biologice de primăvară, vară și toamnă de V.G.Bogorov (1938). Marea Albă este caracterizată de toate cele trei perioade. Durata lor pentru Golful Kandalaksha în 1970 a fost de 48, 92 și, respectiv, 43 de zile. 
Rezultatele care caracterizează valorile absolute ale procesului de producție în diferite perioade ale sezonului de vegetație sunt prezentate în tabel. 5.2. Coeficientul P/Sent a fost calculat ca raportul dintre producția primară și biomasă (exprimată în carbon) pentru Bice TO CTO,JI (5a B OIE bil. 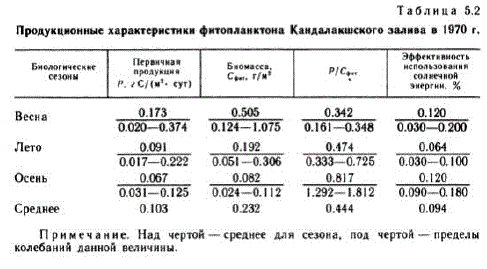
Notă. Deasupra liniei este media pentru sezon, sub linie este intervalul de fluctuații ale unei anumite valori.
Începutul primăverii biologice s-a remarcat în primele zece zile ale lunii aprilie Punctul culminant subglaciar de primăvară (Fedorov, 1970a; Kokin, Koltsova, 1972) durează aproximativ 2 săptămâni și se caracterizează printr-un coeficient P/Cphi stabil. Distribuția econichelor de alge corespunde celui de-al doilea model MacArthur (Kokin și Koltsova, 1972).
Izbucnirea de primăvară a abundenței de biomasă și alge începe sub gheață și atinge un maxim după ce gheața se rupe. Fitoplanctonul este reprezentat de BH La MH JULHETOMOBOTO KOM TIJEKCE: Fragilaria oceanica, Chaetoceros holsa ficus, Nitzschia frigida, Maticula septentrionalia. Cea mai abundentă specie a fost F. oceapisa. Stratul fotic este îmbogățit cu substanțe nutritive și conține 86% din biomasa totală de fitoplancton, care, ca și rata fotosintezei, în această perioadă atinge valoarea maximă pentru întregul sezon de vegetație (vezi Tabelul 5.2). Consumul de alge de către zooplancton (Konoplya, 1973) duce la o scădere a biomasei de alge și la o scădere a competiției alimentare pentru nutrienți, în timp ce distribuția algelor corespunde deja primului model MacArthur.
Maximul de vară al dezvoltării fitoplanctonului (primul), cauzat de creșterea numărului de diatomee, după o scădere a numărului acestora, este „preluat” de algele peridiniu, formând al doilea maxim de vară. De la mijlocul lunii iulie până la sfârșitul lunii august, punctul culminant al verii este stabilit cu un raport stabil P/C-fit. Formele dominante ale verii biologice sunt Sceletopema costalum și speciile din genul Chaetoceros (în principal Ch. compostis). Productia primara in aceasta perioada se caracterizeaza printr-o valoare medie de 0,091 g C/(m", zi), iar biomasa - 0,192 g C/m". Se dezvoltă competiția trofică pentru nutrienți,
Focarul de toamnă al fitoplanctonului este slab exprimat. Se observă de la sfârșitul lunii august până la mijlocul lunii septembrie și creșterea biomasei ee, Rb Ae/y!"b, "pyAhQ. Doar o creștere a producției primare la 0,125 g C/(m", zi) și a coeficientului P/Seite (până la 1812) pe 20 septembrie indică o activare pe termen scurt a fitoplanctonului. În această perioadă, dezvoltarea Dinophyes powegica. , o dominantă iubitoare de frig a toamnei biologice, se observă ( Fedorov, 1969).Perioada de succesiune a toamnei duce ecosistemul la un apogeu pe termen scurt, care durează până la sfârșitul lunii septembrie.O scădere suplimentară a temperaturii și radiației solare, precum si o crestere a stratului de amestec datorita perturbarii saltului de densitate duce la o redistribuire a biomasei fitoplanctonului in profunzime si la predominarea proceselor de distrugere asupra produselor.
În general, un astfel de curs al ciclului de producție a fitoplanctonului poate fi de așteptat în alte zone ale mării. Posibilele diferențe vor fi exprimate în momentul începerii și sfârșitului sezonului de vegetație a fitoplanctonului, precum și în unele diferențe în compoziția calitativă a speciilor de alge dominante (Zhitina, 1981), care vor afecta fără îndoială rezultatele proceselor de producție a fitoplanctonului. în aceste părţi ale zonei de apă.
Datele din studiile ciclului de producție a fitoplanctonului din 1970 au făcut posibilă clarificarea clasificării troficității Mării Albe. Ca o caracteristică suplimentară, a fost utilizată eficiența utilizării energiei solare de către fitoplancton (vezi Tabelul 5.2), care este o caracteristică integrală a stării fitoplanctonului, în funcție de un set de condiții de mediu și neprezentând o legătură directă cu biomasa fitoplanctonului. (Fedorov, Bobrov, 1977).
Dacă comparăm valorile calculate ale principalelor caracteristici de producție ale fitoplanctonului cu datele cunoscute pentru regiunile oceanice cu productivitate diferită (Sorokin, 1973), putem concluziona că Golful Kandalaksha aparține unor zone neproductive cu un nivel de producție de fitoplancton de 0,05-0,10 g. C/(m* - zi) și o eficiență a utilizării energiei solare de 0,04%, caracteristică apelor oligotrofe. În consecință, Golful Kandalaksha, conform clasificării O.I. Koblenz-Mishke (Koblenz-Mishke și colab., 1970), poate fi clasificat ca clasa trofică 1 cu foarte puțin amestec de ape adânci.
Cele mai mici organisme ale coloanei de apă sunt combinate în conceptul de „plancton” (din limba greacă „ planctos„- înălțare, rătăcire). Lumea planctonului este imensă și diversă. Aceasta include organisme care locuiesc în grosimea mărilor, oceanelor, lacurilor și râurilor. Ei trăiesc oriunde există cea mai mică cantitate de apă. Acestea pot fi chiar și cele mai obișnuite bălți, o vază cu flori cu apă stătătoare, fântâni etc.
Comunitatea de plancton este cea mai veche și mai importantă din multe puncte de vedere. Planctonul există de aproximativ 2 miliarde de ani. Au fost primele organisme care au locuit cândva planeta noastră. Organismele plancton au fost primele care au furnizat planeta noastră cu oxigen. Și acum aproximativ 40% din oxigen este produs de plante acvatice, în primul rând planctonice. Planctonul are o mare importanță în echilibrul nutrițional al ecosistemelor acvatice, deoarece multe specii de pești, balene și unele păsări se hrănesc cu ele. Este principala sursă de viață în mări și oceane, lacuri mari și râuri. Impactul planctonului asupra resurselor de apă este atât de mare încât poate afecta chiar compoziția chimică a apelor.
Planctonul include fitoplancton, bacterioplancton și zooplancton. Acestea sunt în principal organisme mici, a căror dimensiune cel mai adesea nu depășește zeci de micrometri pentru alge și câțiva centimetri pentru zooplancton. Cu toate acestea, majoritatea animalelor au dimensiuni semnificativ mai mici. De exemplu, dimensiunea celei mai mari dafnii de apă dulce ajunge la doar 5 mm.
Cu toate acestea, majoritatea oamenilor știu foarte puțin despre plancton, deși numărul de organisme din corpurile de apă este extrem de mare. De exemplu, numărul de bacterii dintr-un centimetru cub de apă ajunge la 5-10 milioane de celule, alge în același volum - zeci până la sute de mii și organisme zooplancton - sute de exemplare. Aceasta este o lume aproape invizibilă. Acest lucru se datorează faptului că majoritatea organismelor cu plancton au dimensiuni foarte mici și pentru a le vizualiza, aveți nevoie de un microscop cu o mărire destul de mare. Organismele care alcătuiesc planctonul plutesc în coloana de apă. Ei nu pot rezista să fie purtati de curenți. Cu toate acestea, acest lucru poate fi discutat doar în termeni generali, deoarece în apa calmă multe organisme planctonice se pot mișca (deși încet) într-o anumită direcție. Algele, care își schimbă flotabilitatea, se mișcă vertical în câțiva metri. Ziua se află în stratul superior de apă bine luminat, iar noaptea coboară cu trei până la patru metri mai adânc, unde sunt mai multe minerale. Zooplanctonul din mări și oceane se ridică în straturile superioare noaptea, unde filtrează algele microscopice, iar dimineața coboară la o adâncime de 300 de metri sau mai mult.
Cine face parte din plancton? Majoritatea organismelor planctonice își petrec întreaga viață în coloana de apă și nu sunt asociate cu substratul solid. Deși etapele de odihnă ale multora dintre ele se stabilesc pe fundul rezervorului în timpul iernii, unde așteaptă condiții nefavorabile. În același timp, printre aceștia se numără și cei care își petrec doar o parte din viață în coloana de apă. Acesta este meroplancton (din limba greacă " meros» - Parte). Se dovedește că larvele multor organisme de fund - arici de mare, stele, stele fragile, viermi, moluște, crabi, corali și altele duc un stil de viață planctonic, sunt purtate de curenți și, în cele din urmă, găsesc locuri pentru un habitat suplimentar, se stabilesc în jos și sunt complet viața nu-l părăsește. Acest lucru se datorează faptului că organismele de fund sunt dezavantajate în comparație cu planctonul, deoarece Se deplasează relativ încet dintr-un loc în altul. Datorită larvelor planctonice, acestea sunt transportate de curenți pe distanțe lungi, la fel cum semințele plantelor terestre sunt purtate de vânt. Ouăle unor pești și larvele lor duc, de asemenea, un stil de viață planctonic.
După cum am observat deja, majoritatea organismelor planctonice sunt adevărate planctere. Se nasc în coloana de apă și acolo mor. Este format din bacterii, alge microscopice, diverse animale (protozoare, rotifere, crustacee, moluște, celenterate etc.).
Organismele planctonice au dezvoltat adaptări care le fac mai ușor să se înalțe în coloana de apă. Acestea sunt tot felul de excrescențe, aplatizarea corpului, incluziuni de gaz și grăsime și un schelet poros. La moluștele planctonice a avut loc reducerea cochiliei. Spre deosebire de organismele bentonice, este foarte subțire și uneori abia vizibilă. Multe organisme planctonice (cum ar fi meduzele) au țesut gelatinos. Toate acestea le permit să-și mențină corpul în coloana de apă fără nicio cheltuială semnificativă de energie.
Multe crustacee planctonice suferă migrații verticale. Noaptea, se ridică la suprafață, unde mănâncă alge, iar mai aproape de zori coboară la o adâncime de câteva sute de metri. Acolo, în întuneric, se ascund de pești, care îi mănâncă cu plăcere. În plus, temperatura scăzută reduce metabolismul și, în consecință, cheltuielile de energie pentru menținerea funcțiilor vitale. La adâncimi mari, densitatea apei este mai mare decât la suprafață, iar organismele se află într-o stare de flotabilitate neutră. Acest lucru le permite să rămână în coloana de apă fără costuri. Fitoplanctonul locuiește în principal în straturile de suprafață ale apei în care pătrunde lumina soarelui. La urma urmei, algele, ca și plantele terestre, au nevoie de lumină pentru a se dezvolta. În mări trăiesc la o adâncime de 50-100 m, iar în corpurile de apă dulce - până la 10-20 de metri, ceea ce se datorează transparenței diferite a acestor corpuri de apă.
În oceane, adâncimea habitatului algelor este cea mai subțire peliculă a unei grosimi uriașe de apă. Cu toate acestea, în ciuda acestui fapt, algele microscopice sunt hrana principală pentru toate organismele acvatice. După cum sa menționat deja, dimensiunea lor nu depășește câteva zeci de micrometri. Doar dimensiunea coloniilor ajunge la sute de micrometri. Crustaceele se hrănesc cu aceste alge. Dintre aceștia, suntem cel mai familiarizați cu krill, care include în principal crustacee euphausiide de până la 1,5 cm în dimensiune.Crustaceele sunt mâncate de peștii planctivori, iar ei, la rândul lor, sunt mâncați de pești mai mari și mai răpitori. Balenele se hrănesc cu krill și le filtrează în cantități uriașe. Astfel, 5 milioane dintre aceste crustacee au fost găsite în stomacul unei balene albastre de 26 m lungime.
Planctonul fitoplancton marin este format în principal din diatomee și piridinii. Diatomeele domină în apele polare și subpolare ale mării (oceanelor). Sunt atât de multe, încât scheletele de siliciu formează sedimente de fund după ce mor. Suflul de diatomee acoperă cea mai mare parte a fundului mărilor reci. Ele apar la adâncimi de aproximativ 4000 m sau mai mult și constau în principal din valve de diatomee mari. Cojile mici se dizolvă de obicei înainte de a ajunge la fund. Mineralul diatomit este un produs al diatomeelor. Numărul de valve din diatomee în unele zone ale oceanului ajunge la 100-400 de milioane la 1 gram de mâl. Scurgetele de diatomee se transformă în cele din urmă în roci sedimentare, din care se formează „pământul de diatomee” sau mineralul diatomit. Este format din cochilii poroase de silex minuscule și este folosit ca material filtrant sau sorbant. Acest mineral este folosit pentru a face dinamită.
În 1866-1876. Chimistul și antreprenorul suedez Alfred Nobel căuta modalități și mijloace de a produce un exploziv puternic. Nitroglicerina este un exploziv foarte eficient, dar explodează spontan cu mici șocuri. După ce a stabilit că pentru a preveni exploziile este suficient să se înmoaie pământul de diatomee în nitroglicerină lichidă, Nobel a creat un exploziv sigur - dinamita. Astfel, îmbogățirea lui Nobel și celebrele „Premii Nobel” stabilite prin testamentul său își datorează existența celor mai mici diatomee.
Apele calde ale tropicelor se caracterizează printr-o diversitate mai mare a speciilor în comparație cu fitoplanctonul mărilor arctice. Cele mai diverse alge de aici sunt peridinea. Cocolitoforele calcaroase flagelate și silicoflagelatele sunt răspândite în planctonul marin. Cocolitoforele locuiesc în principal în apele tropicale. Mâlurile calcaroase, inclusiv scheletele cocolitoforelor, sunt răspândite în Oceanul Mondial. Cel mai adesea se găsesc în Oceanul Atlantic, unde acoperă mai mult de 2/3 din suprafața fundului. Cu toate acestea, nămolurile conțin cantități mari de cochilii de foraminifere aparținând zooplanctonului.
Observațiile vizuale ale apelor mării sau oceanului fac posibilă determinarea cu ușurință a distribuției planctonului după culoarea apei. Albastrul și transparența apelor indică sărăcia vieții; într-o astfel de apă practic nu există nimeni care să reflecte lumina în afară de apa însăși. Albastrul este culoarea deserturilor maritime, unde organismele plutitoare sunt foarte rar întâlnite. Culoarea verde este un indicator inconfundabil al vegetației. Prin urmare, atunci când pescarii întâlnesc apă verde, ei știu că straturile de suprafață sunt bogate în vegetație, iar acolo unde sunt multe alge, există întotdeauna o abundență de animale care se hrănesc cu ea. Fitoplanctonul este numit pe bună dreptate pășunea mării. Algele microscopice sunt hrana principală a unui număr mare de locuitori ai oceanului.
Culoarea verde închis a apei indică prezența unei mase mari de plancton. Nuanțele de apă indică prezența anumitor organisme planctonice. Acest lucru este foarte important pentru pescari, deoarece natura planctonului determină tipul de pește care trăiește în zonă. Un pescar experimentat poate detecta cele mai subtile nuante de culoare din apa de mare. În funcție de faptul că pescuiește în apă „verde”, „galbenă” sau „roșie”, un „ochi cu experiență” poate prezice cu un grad rezonabil de probabilitate natura și mărimea capturii.
În corpurile de apă dulce predomină algele albastru-verzi, verzi, diatomee și dinofite. Dezvoltarea abundentă a fitoplanctonului (așa-numita „înflorire” a apei) schimbă culoarea și transparența apei. În corpurile de apă dulce se observă cel mai adesea înfloririle albastru-verzui, iar în mări se observă înflorirea peridinei. Substanțele toxice pe care le eliberează reduc calitatea apei, ceea ce duce la otrăvirea animalelor și a oamenilor, iar în mări provoacă moartea în masă a peștilor și a altor organisme.
Culoarea apei în anumite zone sau mări este uneori atât de caracteristică încât mările și-au luat numele de la culoarea apei. De exemplu, culoarea deosebită a Mării Roșii este cauzată de prezența algelor albastre-verzi Trichodesmium ( Trichodesmium egythraeum), care are un pigment care conferă apei o nuanță brun-roșcată; sau Crimson Sea - fostul nume al Golfului California.
Unele dinoflagelate de plante (de exemplu, Gonyaulax și Gymnodinium) dau apei o culoare deosebită.În apele tropicale și temperate calde, aceste creaturi se reproduc uneori atât de repede încât marea devine roșie. Pescarii numesc acest fenomen „marea roșie”. Acumulările uriașe de dinoflagelate (până la 6 milioane de celule într-un litru de apă) sunt extrem de otrăvitoare, așa că în timpul „mareei roșii” multe organisme mor. Aceste alge nu sunt doar otrăvitoare în sine; ei eliberează substanțe toxice, care apoi se acumulează în organismele care mănâncă dinoflagelate. Orice creatură, fie că este un pește, o pasăre sau o persoană, care mănâncă un astfel de organism devine periculos otrăvită. Din fericire, fenomenul mareei roșii este local și nu se întâmplă des.
Apele mărilor sunt colorate nu numai de prezența algelor, ci și de zooplancton. Majoritatea euphausiidelor sunt transparente și incolore, dar unele sunt de culoare roșu aprins. Astfel de euphausiide trăiesc în emisferele nordice și sudice mai reci și uneori se acumulează într-un asemenea număr încât întreaga mare devine roșie.
Colorarea apei este dată nu numai de alge planctonice microscopice, ci și de diferite particule de origine organică și anorganică. După ploile abundente, râurile aduc o mulțime de particule minerale, motiv pentru care apa capătă nuanțe diferite. Astfel, particulele de argilă aduse de râul Galben dau Mării Galbene o nuanță corespunzătoare. Râul Galben (din chineză - Râul Galben) și-a primit numele datorită turbidității sale. Multe râuri și lacuri conțin atât de mulți compuși humici încât apele lor devin închise - maro și chiar negre. De aici și numele multora dintre ei: Rio Negro - în America de Sud, Black Volta, Niger - în Africa. Multe dintre râurile și lacurile noastre (și orașele situate pe ele) sunt numite „negre” din cauza culorii apei.
În corpurile de apă dulce, colorarea apei datorită dezvoltării algelor apare mai des și mai intens. Dezvoltarea masivă a algelor provoacă fenomenul de „înflorire” a corpurilor de apă. În funcție de compoziția fitoplanctonului, apa este colorată în diferite culori: de la algele verzi Eudorina, Pandorina, Volvox - verde; din diatomee Asterionella, Tabellaria, Fragilaria – culoare brun-gălbui; din flagelate Dinobryon – verzui, Euglena – verde, Synura – maro, Trachelomonas – maro-gălbui; din dinofit Ceratium - galben-brun.
Biomasa totală a fitoplanctonului este mică în comparație cu biomasa zooplanctonului care se hrănește cu acesta (respectiv, 1,5 miliarde de tone și peste 20 de miliarde de tone). Cu toate acestea, datorită reproducerii rapide a algelor, producția (recolta) acestora în Oceanul Mondial este de aproape 10 ori mai mare decât producția totală a întregii populații vii a oceanului. Dezvoltarea fitoplanctonului depinde în mare măsură de conținutul de substanțe minerale din apele de suprafață, cum ar fi fosfații, compușii de azot și altele. Prin urmare, în mări, algele se dezvoltă cel mai abundent în zonele cu ape adânci în creștere, bogate în minerale. În corpurile de apă dulce, afluxul de îngrășăminte minerale spălate de pe câmpuri și diverse ape uzate menajere și agricole duce la dezvoltarea masivă a algelor, care afectează negativ calitatea apei. Algele microscopice se hrănesc cu organisme planctonice mici, care la rândul lor servesc drept hrană pentru organisme mai mari și pești. Prin urmare, în zonele cu cea mai mare dezvoltare a fitoplanctonului există o mulțime de zooplancton și pești.
Rolul bacteriilor în plancton este mare. Ele mineralizează compușii organici (inclusiv diverși poluanți) ai corpurilor de apă și îi reintroduc în ciclul biotic. Bacteriile în sine sunt hrană pentru multe organisme zooplancton. Numărul de bacterii planctonice din mări și corpuri de apă curată nu depășește 1 milion de celule într-un mililitru de apă (un centimetru cub). În majoritatea corpurilor de apă dulce, numărul lor variază între 3-10 milioane de celule într-un mililitru de apă.
A.P. Sadcikov,
Profesor la Universitatea de Stat din Moscova, numit după M.V. Lomonosov, Societatea Oamenilor de Științe Naturale din Moscova
(http://www.moip.msu.ru)
TI-A PLACUT MATERIALUL? ABONAȚI-VĂ LA NEWSLETTERUL NOSTRU DE EMAIL:
În fiecare luni, miercuri și vineri vă vom trimite un rezumat prin e-mail cu cele mai interesante materiale de pe site-ul nostru.
Sleepsova E.V. 1Savvina S.R. 1
Vakhrusheva A.V. 1 Ivanova A.P. 2
1 Instituție bugetară educațională municipală școala secundară nr. 21 din districtul urban „Orașul Iakutsk”
2 IBPK SB RAS
Textul lucrării este postat fără imagini și formule.
Versiunea completă a lucrării este disponibilă în fila „Fișiere de lucru” în format PDF
INTRODUCERE
Lacurile orașului sunt de mare importanță ecologică pentru oraș, deoarece sunt o sursă de alimentare cu apă menajeră, un habitat pentru pești și, de asemenea, un loc de recreere pentru oameni. Datorită deteriorării situației ecologice în mediul urban, este necesar să se efectueze observații ale compoziției hidrobiologice a mediului acvatic.
Scopul lucrării este de a identifica compoziția de specii a fitoplanctonului pentru a evalua starea actuală a rezervorului și de a elabora recomandări pentru îmbunătățirea situației ecologice a lacului Soldatskoye.
Ipoteza cercetării: Se poate presupune că punerea în aplicare la timp a măsurilor de securitate va face posibilă conservarea lacului Soldatskoye și, de asemenea, va face posibilă crearea unei zone de securitate în interiorul orașului.
Pentru atingerea acestui obiectiv au fost stabilite următoarele: sarcini :
- Determinați compoziția fitoplanctonului.
- Identificați specii-indicatori ai saprobității.
- Evaluați starea actuală a fitoplanctonului și elaborați recomandări pentru îmbunătățirea stării rezervorului.
Semnificație științifică și practică. Materialele acestei lucrări pot servi drept bază pentru monitorizarea mediului înconjurător al corpurilor de apă urbane și pot fi folosite și ca materiale de propagandă dedicate problemelor de îmbunătățire a infrastructurilor urbane și problemelor de respect pentru elementele naturii.
Capitolul 1. Fitoplanctonul rezervoarelor
Un rezervor ca ecosistem este un complex al tuturor organismelor și elementelor neînsuflețite, ca urmare a interacțiunii căreia fluxul de energie într-un loc dat creează o structură stabilă și o circulație a substanțelor (Lasukov, 2009).
Componentele principale ale ecosistemului acvatic:
1) energie primită de la Soare;
2) factorii climatici și fizici;
3) compuși anorganici;
4) compuși organici;
5) producători de compuși organici, sau producători(din lat. producentis- creând) - plante înrădăcinate, plutitoare libere și alge minuscule (fitoplancton, din greacă. phytos- plantă, plancton- rătăcire, înălțare);
6) consumatorii primari, sau consumatorii primari(din lat. consum- consumă), hrănirea cu plante - zooplancton (plancton animal), moluște, larve, mormoloci;
7) consumatori secundari, sau consumatori secundari- insecte și pești răpitori;
8) detritus(din lat. detritus- uzate) - produse de degradare și descompunere a organismelor;
9) distrugători, distrugători, descompunetori(din lat. reduceti s - întoarcere, restabilire), detritivore(din greaca phagos- devorator), saprotrofe(din greaca sapros- putred și trofeu e - nutriție) - bacterii și ciuperci de fund, larve, moluște, viermi.
O componentă importantă a ecosistemelor lacustre sunt algele planctonului (fitoplanctonul). Fitoplanctonul este principalul producător de materie organică din corpurile de apă, datorită căruia există majoritatea animalelor acvatice. Sunt sensibili la schimbările de mediu și joacă un rol important de bioindicator. Fitoplanctonul influențează dezvoltarea nevertebratelor în plancton (zooplancton), care, la rândul său, este un filtru natural și sursă de hrană pentru pești.
Fitoplanctonul include alge protococice, diatomee, dinoflagelate, cocolitofore și alte alge unicelulare (adesea coloniale), precum și cianobacteriile. Trăiește în zona fotică a corpurilor de apă, locuind în coloana de apă. Abundența fitoplanctonului în diferite părți ale corpurilor de apă depinde de cantitatea de nutrienți necesari acestuia în straturile de suprafață. Factorii limitanți în acest sens sunt în principal fosfații, compușii de azot, iar pentru unele organisme (diatomee, flagelati de silice) și compușii de siliciu. Deoarece animalele planctonice mici se hrănesc cu fitoplancton și servesc drept hrană pentru cele mai mari, zonele cu cea mai mare dezvoltare a fitoplanctonului sunt, de asemenea, caracterizate de o abundență de zooplancton și necton. Scurgerea râului are o semnificație mult mai mică și doar locală în îmbogățirea apelor de suprafață cu substanțe nutritive. Dezvoltarea fitoplanctonului depinde și de intensitatea luminii, care în apele reci și temperate determină sezonalitate în dezvoltarea planctonului. În timpul iernii, în ciuda abundenței de nutrienți transportați în straturile de suprafață ca urmare a amestecului de iarnă a apelor, există puțin fitoplancton din cauza lipsei de lumină. Primavara incepe dezvoltarea rapida a fitoplanctonului, urmata de zooplancton. Pe măsură ce fitoplanctonul folosește nutrienți, precum și datorită consumului său de către animale, cantitatea de fitoplancton scade din nou. La tropice, compoziția și cantitatea de plancton sunt mai mult sau mai puțin constante pe tot parcursul anului. Dezvoltarea abundentă a fitoplanctonului duce la așa-numita „înflorire” a apei, schimbându-i culoarea și reducând transparența apei. Când unele specii înfloresc, substanțele toxice sunt eliberate în apă, care pot provoca decese în masă ale animalelor planctonice și nectonice, precum și reacții alergice ale pielii, conjunctivită și tulburări gastro-intestinale la om.
După mărime planctonul este împărțit în:
1) megaloplancton (megalos - imens) - care include organisme mai mari de 20 cm;
2) macroplancton (makros - mare) - 2-20 cm;
3) mezoplancton (mezo - medie) -0,2-20 mm;
4) microplancton (mikros - mic) - 20-200 microni;
5) nanoplancton (nanos - pitic) - 2-20 microni;
6) picoplancton - 0,2-2 um;
7) femtoplancton (virusuri oceanice) -< 0,2 мкм.
Cu toate acestea, limitele acestor grupuri de mărime nu sunt în general acceptate. Multe organisme de plancton au dezvoltat adaptări care facilitează plutirea în apă: reducerea masei specifice a corpului (incluziuni de gaze și grăsimi, saturație cu apă și țesut gelatinos, subțierea și porozitatea scheletului) și creșterea suprafeței sale specifice (complex, adesea excrescente foarte ramificate, corp turtit).
Biomasa fitoplanctonului variază în diferite corpuri de apă și regiunile acestora, precum și în diferite anotimpuri. În lacurile orașului Yakutsk, biomasa variază între 0,255-3,713 mg/l (Ivanova, 2000). Odată cu adâncimea, fitoplanctonul devine mai puțin divers și cantitatea lui scade rapid, cu valori maxime la o adâncime de 1-2 transparență. Transparența apei în hidrologie și oceanologie este raportul dintre intensitatea luminii care trece printr-un strat de apă și intensitatea luminii care intră în apă. Transparența apei este o valoare care indică indirect cantitatea de particule și coloizi în suspensie din apă. Producția anuală de fitoplancton în Oceanul Mondial este de 550 de miliarde de tone (conform estimărilor oceanologului sovietic V.G. Bogorov), ceea ce este de aproape 10 ori mai mare decât producția totală a întregii populații de animale a oceanului.
Fitoplanctonul, în special lacurile, în procesul de formare și dezvoltare poate suferi o serie de modificări datorită naturii habitatului ecologic: caracteristicile locației și morfometriei rezervorului, compoziția chimică specifică a apei, fluctuațiile de nivel, alimentarea. a nutrientilor din apa, rezultatul activitatii economice umane etc. Aceasta duce la inlocuirea unor tipuri de alge cu altele, mai specializate. În general, diversitatea și compoziția generală a speciilor de fitoplancton pot servi ca indicatori ecologici buni. Studiile comparative ale modelelor de distribuție ale compoziției, structurii și productivității fitoplanctonului în corpurile de apă ale diferitelor zone naturale devin din ce în ce mai relevante, creând baza pentru dezvoltarea stării lor trofice și pentru prezicerea schimbărilor de mediu în ecosistemele acvatice sub influența încărcăturii antropice (Ermolaev). , 1989).
Taxonomia modernă a algelor include 13 diviziuni:
Cianoprocariote- alge albastre-verzi (cianobacterii);
Euglenophyta- alge euglene;
Chrysophyta- alge crizofite;
Xanthophyta- alge galben-verzui;
Eustigmatophyta- alge eustigmatoide;
Bacillariophyta- diatomee;
Dinophyta- alge dinofite;
cryptophyta- alge criptofite;
Raphydophyta- alge rafidofite;
Rhodophyta- alge roșii;
Phaeophyta- alge brune;
Chlorophyta- alge verzi;
Streptophyta- alge streptofite.
Capitolul 2. Materiale şi metode de cercetare hidrobiologică
Pentru a identifica compoziția actuală a fitoplanctonului din Lacul Soldatskoye, prelevarea de probe a fost efectuată în vara anului 2017 (23 mai, 21 iunie, 12 iulie). Probele de apă au fost prelevate de la orizontul de suprafață al lacului de acumulare din zona litoralului. Prelevarea a fost efectuată la două stații: în apropierea pieței Ryzhikov (locul 1) și lângă restaurantul Panda (locul 2). Informațiile despre punctele de prelevare sunt date în Tabelul 1 (fotografiile 1 și 2).
tabelul 1
Puncte de prelevare a apei pentru studiile fitoplanctonului
|
Proba nr. |
Data |
Numele punctului de prelevare |
Calitate/număr |
Volumul de apă filtrată (l) |
|||
|
Parcela nr. 1 (lângă Piața Ryzhikov) |
|||||||
|
Secțiunea nr. 2 |
|||||||
|
Parcela nr. 1 (lângă Piața Ryzhikov) |
|||||||
|
Secțiunea nr. 2 (lângă restaurantul Panda and Crane |
|||||||
|
Parcela nr. 1 (lângă Piața Ryzhikov) |
|||||||
|
Secțiunea nr. 2 (lângă restaurantul Panda and Crane |
|||||||
|
FOTO 1. Eșantionare la locul 1 (lângă Piața Ryzhikov) |
FOTO 2. Eșantionare în zona 2 (lângă cafeneaua Panda) |
||||||
Alegerea metodei de prelevare a fitoplanctonului depinde de tipul rezervorului, gradul de dezvoltare al algelor, obiectivele cercetării, instrumentele disponibile, echipamentele etc. Se folosesc diverse metode de preconcentrare a microorganismelor. O astfel de metodă este filtrarea apei prin rețele de plancton.
Rețeaua de plancton constă dintr-un inel de alamă și o pungă conică cusute la ea dintr-o sită de moară de mătase sau nailon nr. 30 (Fig. 1). O diagramă a modelului unui con de plasă pentru o plasă de plancton este prezentată în Figura 2. Ieșirea îngustă a pungii în formă de con este atașată strâns de o cană având un tub de evacuare închis cu un robinet sau clemă Mohr. În corpuri mici de apă, probele de plancton pot fi colectate de pe țărm, aruncând o plasă pe o frânghie subțire în apă și trăgând-o cu grijă. În corpurile mari de apă, probele de plancton sunt prelevate dintr-o barcă. Se recomandă tragerea plasei de plancton în spatele unei bărci în mișcare timp de 5-10 minute. După ce s-a terminat de colectat probele de plancton, plasa de plancton este clătită coborând-o de mai multe ori în apă până la inelul superior pentru a spăla algele care au rămas pe suprafața interioară a plasei. Proba concentrată, situată în cupa plasei de plancton, se toarnă prin tubul de evacuare într-un borcan sau o sticlă curată pregătită în prealabil. Înainte de a începe și după finalizarea recoltării probei, este necesar să clătiți bine plasa, după terminarea lucrării, uscați-o și puneți-o într-o cutie specială. Aceste mostre pot fi studiate într-o stare vie și fixă. Pentru depozitare pe termen lung, adăugați o soluție de formaldehidă 40% la probă la o rată de 2-3 picături la 10 ml.
Pentru înregistrarea cantitativă a fitoplanctonului se prelevează probe de un anumit volum. În aceste scopuri pot fi utilizate și taxele de rețea, sub rezerva contabilizării obligatorii a cantității de apă filtrată prin rețea și a volumului probei colectate. De obicei, eșantionarea pentru contabilizarea cantitativă a fitoplanctonului se realizează cu dispozitive speciale - batometre de diferite modele. Când se studiază fitoplanctonul în straturile de suprafață de apă, probele sunt prelevate prin introducerea apei într-un vas de un anumit volum. În rezervoarele cu fitoplancton sărac, este indicat să se preleveze probe de cel puțin 1 litru în paralel cu colecții de plasă care permit captarea obiectelor mici, relativ mari.
În rezervoarele cu fitoplancton bogat, volumul unei probe cantitative poate fi redus la 0,5 sau chiar 0,25 l (de exemplu, atunci când apa „înflorește”). Am filtrat 10 litri de apă folosind o găleată printr-o rețea Apstein și am fixat, de asemenea, cu formaldehidă 40%.
Etichetarea și ținerea unui jurnal de teren
Toate probele colectate sunt etichetate. Pe etichete, cu creion, se indică numărul probei, rezervorul, numărul stației, orizontul de prelevare, volumul de apă filtrată, dacă această probă a fost prelevată pentru analiza cantitativă, data și numele colectorului. Eticheta este plasată în recipientul care conține proba. Aceleași date sunt introduse în jurnalul de teren; în plus, ele indică temperatura aerului și a apei, un desen schematic al rezervorului indicând stațiile de prelevare, o descriere detaliată a rezervorului studiat și a vegetației acvatice superioare și alte informații (vânt, tulburare etc.).
Metode de studiu calitativ al materialului
Materialul colectat este examinat preliminar la microscop în stare de viață în ziua recoltării pentru a se observa starea calitativă a algelor înainte de apariția modificărilor cauzate de depozitarea materialului viu sau de fixarea probelor (formarea celulelor reproductive, a coloniilor). , pierderea flagelilor și a motilității etc.). Ulterior, materialul colectat continuă să fie studiat în stare fixă. Algele au fost studiate folosind diferite mărci de microscoape ușoare folosind diferite sisteme oculare și obiective în lumină transmisă, urmând regulile uzuale ale microscopiei.
Pentru studiul microscopic al algelor se prepară preparate: o picătură din lichidul studiat este plasată pe o lamă de sticlă și acoperită cu o sticlă de acoperire. Când studiați medicamentul pentru o lungă perioadă de timp, lichidul de sub capacul de sticlă se usucă treptat, așa că ar trebui adăugat. Pentru a reduce evaporarea, se aplică un strat subțire de parafină de-a lungul marginilor lamei.
Metode pentru măsurarea dimensiunii algelor și determinarea valorii diviziunii unui micrometru pentru ocular
Când se studiază compoziția speciilor de alge, se măsoară dimensiunea acestora, ceea ce este o caracteristică importantă de diagnosticare. Pentru a măsura obiectele microscopice, se folosește un micrometru pentru ocular cu o riglă de măsurare. Valoarea diviziunii micrometrului ocularului este determinată folosind un micrometru obiect individual pentru fiecare microscop și obiectiv. Obiectul micrometru este o lamă de sticlă cu o riglă aplicată, a cărei lungime este de 1 mm. Rigla este împărțită în 100 de părți, astfel încât fiecare parte să fie egală cu 0,01 mm sau 10 µm. Pentru a afla cu ce este egală o diviziune a riglei ocularului la o mărire dată, este necesar să se stabilească o corespondență între diviziunile (liniile) riglei ocularului de măsurare și obiectul micrometru. De exemplu: 10 diviziuni ale micrometrului pentru ocular coincid cu 5 diviziuni ale micrometrului obiect (adică egale cu 0,05 mm). Prin urmare, o diviziune a riglei ocularului este egală cu 0,05 mm: 10 = 0,005 mm = 5 μ (µm). Acest calcul trebuie făcut pentru fiecare lentilă de 3-4 ori pentru a obține o valoare a diviziunii mai precisă.
La studierea dimensiunilor liniare ale algelor este indicat să se măsoare cât mai multe exemplare (10-100) cu prelucrarea statistică ulterioară a datelor obţinute. La identificarea algelor, trebuie să se obțină precizie. Când studiați materialul original, este necesar să notați orice abateri, chiar și minore, de la diagnostice în dimensiune, formă și alte caracteristici morfologice și să le înregistrați în descrierile, desenele și microfotografiile dumneavoastră.
În practica cercetării algologice, microscopia electronică cu transmisie și scanare este din ce în ce mai utilizată. Metodele pentru prepararea și studiul medicamentelor sunt descrise în literatura de specialitate.
Metode de numărare cantitativă a algelor
Pot fi numărate numai probe cantitative de fitoplancton. Datele privind numărul de alge reprezintă punctul de plecare pentru determinarea biomasei acestora și recalcularea altor indicatori cantitativi (conținut de pigmenți, proteine, grăsimi, carbohidrați, vitamine, acizi nucleici, elemente de cenușă, rata respirației, fotosinteza etc.) pe celulă sau pe fiecare. unitate de biomasă. Numărul poate fi exprimat în numărul de celule, cenobii, colonii, secțiuni de fire de o anumită lungime etc.
Numărarea numărului de alge se efectuează pe pahare de numărare speciale (grafice în benzi și pătrate), pe suprafața cărora se aplică o picătură de apă dintr-o probă de testare bine amestecată cu o pipetă de ștampilă de un anumit volum (în cea mai mare parte 0,1 cm). 3). Pentru a număra numărul de alge, se folosesc și camere de numărare ale Nageotte cu un volum de 0,01 cm 3 și „Uchinskaya” (0,02 cm 3). În plus, puteți utiliza camere folosite pentru numărarea celulelor sanguine - Goryaev, cu un volum de 0,9 mm 3, Fuchs-Rosenthal, etc. Când utilizați camere Goryaev și Fuchs-Rosenthal, geamul de acoperire este frecat cu grijă de suprafețele laterale ale se numără lama de sticlă până când apar inele Newton, apoi se umple camera cu o picătură din proba de testat folosind o pipetă. În funcție de numărul de organisme din proba de testat, pot fi numărate fie toate, fie o parte din urmele (pătratele) de pe suprafața paharului de numărare. Este necesar să se efectueze numărări repetate a mai multor (cel puțin trei) picături din aceeași probă, folosind de fiecare dată o pipetă pentru a selecta o probă pentru numărare, după agitarea temeinică a probei.
Când se studiază probe cantitative de fitoplancton, numărul de organisme per 1 litru de apă este recalculat folosind formula
N=¾¾¾, unde
N - număr (celule/l),
n este numărul mediu de celule numărate în cameră,
V 1 - volumul de apă filtrată (l),
V 2 - volumul probei (ml),
V 3 - volumul camerei (ml).
Conținutul cantitativ al algelor din eșantioane este reflectat cel mai pe deplin de indicatorii biomasei lor, care sunt determinați prin numărare-volum, greutate, volum și diverse metode chimice (radiocarbon, clorofilă etc.).
Pentru a determina biomasa folosind metoda numărării volumului, este necesar să existe date despre numărul acestora în fiecare probă specifică pentru fiecare specie separat și volumele lor medii (pentru fiecare specie din fiecare probă specifică). Există diferite metode pentru a determina volumul corporal al algelor. Cea mai exactă este metoda stereometrică, în care corpul algelor este echivalat cu un corp geometric sau o combinație de astfel de corpuri, după care volumele acestora sunt calculate folosind formule cunoscute în geometrie pe baza dimensiunilor liniare ale unor organisme specifice. Uneori, folosesc volume medii corporale gata făcute, calculate anterior pentru diferite tipuri de alge, care sunt date în lucrările multor autori. Biomasa este calculată pentru fiecare specie separat, iar apoi datele sunt însumate. Metoda de numărare a volumului pentru determinarea biomasei este utilizată pe scară largă în practica cercetării hidrobiologice atunci când se studiază relațiile cantitative ale diferitelor componente ale biocenozelor, modelele de distribuție a algelor în diferite biotopi ale aceluiași rezervor sau în rezervoare diferite, sezoniere și pe termen lung. dinamica dezvoltării algelor etc.
Biomasa algelor este determinată conform metodei general acceptate (Makarova et al., 1970) prin echivalarea celulelor individuale cu figuri geometrice (Fig. 3) folosind tabele standard (Kuzmin, 1984) iar biomasa este calculată folosind formula:
N este numărul de celule în 1 l (celule/l);
W - greutatea celulei (mg).
În lipsa tabelelor standard, calculăm volumul și greutatea celulei (W) folosind formule geometrice (Fig. 3): pentru un cilindru cu o înălțime foarte mică (B) V = πr 2 h; un cilindru cu o elipsă la bază (A)
V = πabh; cubul V = l 3; paralelipiped V = abc;
bila V = - πr 3 ; conul V = - πr 2 h; elipsoid V = - πabc;
(c + 2b)ah (c + 2b)ah
pană V = ————; 2 pene V = ————
Orice ecuație cu cifre este condiționată, astfel încât erorile sunt posibile atât în direcția de creștere, cât și de scădere a volumului „adevărat” al celulei. Având în vedere acest lucru, este necesar să echivalăm o celulă de fiecare tip, pe cât posibil, cu figura geometrică care se potrivește cel mai mult cu volumul real al unei celule date. După ce calculăm volumul folosind formula, trebuie să înmulțiți volumul rezultat cu 10 -9. Greutatea se măsoară în mg. Pentru a determina mai precis biomasa fitoplanctonului, este necesar să se țină cont de mucusul din jurul celulei, precum și de grosimea cochiliei în diatomee.
Cu dezvoltarea intensivă a algelor, puteți utiliza metoda greutății. În acest caz, proba de testat este filtrată printr-un filtru de hârtie pre-uscat și cântărit (în paralel, apa distilată este filtrată prin filtre de control). Apoi filtrele sunt cântărite și uscate într-un cuptor la 100ºC până la greutate constantă. Pe baza datelor obținute se calculează greutățile uscate și umede ale sedimentului. Ulterior, prin arderea filtrelor într-un cuptor cu mufă, se poate determina conținutul de substanțe organice din sediment. Dezavantajul acestei metode este că dă o idee doar asupra masei totale a tuturor substanțelor organice, organismelor vii și impurităților nevii, de origine animală și vegetală, suspendate în probă. Contribuția reprezentanților taxonilor individuali la această masă totală poate fi exprimată doar aproximativ în fracții de masă după calcularea raportului lor la microscop în mai multe câmpuri de vedere. Cea mai completă imagine a biomasei algelor poate fi obținută prin combinarea mai multor metode de cercetare diferite.
Metoda de determinare a frecvenței
La procesarea calitativă a probelor, este de dorit să se determine frecvența de apariție a speciilor individuale, folosind simboluri pentru aceasta. Există diferite scale pentru frecvența de apariție a algelor:
Frecvența de apariție a speciei (h) în funcție de scara Levander (Levander, 1915) și Ostelfeld (Ostenfeld, 1913) așa cum a fost modificată de Kuzmin (Kuzmin, 1976) are o expresie numerică de la 1 la 6:
rr - foarte rar (de la 1 la 10 mii celule/l) - 1;
r - rar (de la 10 mii celule/l la 100 mii celule/l) - 2;
rc - adesea (de la 100 de mii de celule/l la 1 milion de celule/l) - 3;
s - adesea (de la 1 milion de celule/l la 10 milioane de celule/l) - 4;
ss - foarte des (de la 10 milioane celule/l la 100 milioane celule/l) - 5;
sss - masă, „înflorire” (de la 100 de milioane de celule/l și mai mult) - 6.
Frecvența de apariție a speciei (h) conform scalei Starmach (Starmach, 1955):
Foarte rar (specia nu este prezentă în fiecare preparat);
1 - singur (1-6 exemplare în pregătire);
2 - puține (7-16 exemplare în pregătire);
3 - destul de mult (17-30 de exemplare în pregătire);
4 - multe (31-50 de exemplare în pregătire);
5 - foarte multe, predominanță absolută (peste 50 de exemplare în pregătire).
Utilizarea algelor pentru analiza biologică a apei
Analiza biologică a apei, împreună cu alte metode, este utilizată pentru a evalua starea corpurilor de apă și pentru a monitoriza calitatea apei. Algele, datorită proprietăților stenotopice ale multor specii și sensibilității lor ridicate la condițiile de mediu, joacă un rol important în analiza biologică a apei. Structura fitoplanctonului este foarte sensibilă la condițiile de mediu. Alături de numărul, biomasa și abundența speciilor, indicii de diversitate a speciilor și indicii de informații pot fi promițători ca indicatori ai poluării apei.
Calitatea sau gradul de poluare a apei pe baza compoziţiei algelor se evaluează în două moduri: a) prin organisme indicator; b) pe baza rezultatelor comparării structurii comunităţii în zone cu grade diferite de poluare şi în zona de control. În primul caz, pe baza prezenței sau absenței speciilor sau grupurilor indicator și a cantității lor relative, folosind sisteme pre-dezvoltate de organisme indicator, un rezervor sau zona sa este atribuită unei anumite clase de ape. În al doilea caz, concluzia se face pe baza rezultatelor comparării compoziției algelor la diferite stații sau secțiuni ale rezervorului, care sunt supuse la diferite grade de poluare.
În algologie se folosește un sistem de saprobare a apei, care se apreciază prin gradul de contaminare a acestora cu substanțe organice și produsele lor de degradare. Sistemul de determinare a saprobității, propus în 1908 de R. Kolkwitz și M. Marsson, și modificările sale ulterioare au primit cea mai mare recunoaștere. Acești autori credeau că descompunerea materiei organice conținute în apele reziduale se face treptat. În acest sens, corpurile de apă sau zonele lor, în funcție de gradul de poluare cu substanțe organice, se împart în poli-, mezo- și oligoaprobice.
În zona polisaprobică, situată aproape de locul de evacuare a apelor uzate, descompunerea proteinelor și carbohidraților are loc în condiții aerobe. Această zonă se caracterizează prin absența aproape completă a oxigenului liber, prezența proteinelor necompuse în apă, cantități semnificative de hidrogen sulfurat și dioxid de carbon și natura reductivă a proceselor biochimice. Numărul de specii de alge capabile să se dezvolte în această zonă este relativ mic, dar se găsesc în cantități mari.
În zona mezosaprobică, poluarea este mai puțin pronunțată: nu există proteine necompuse, există puțină hidrogen sulfurat și dioxid de carbon, oxigenul este prezent în cantități notabile, dar apa conține și compuși de azot slab oxidați precum amoniacul, aminoacizii și amidoacizii. Zona mezosaprobică este împărțită în subzone α- și β-mezoaprobice. Primul conține amoniac, aminoacizi și amidoacizi, dar conține deja oxigen. În această zonă se găsesc alge albastre-verzi din genurile Oscillatoria și Formidium. Mineralizarea materiei organice are loc în principal din cauza oxidării aerobe, în special a bacteriilor. Următoarea zonă mezosaprobică este caracterizată prin prezența amoniacului și a produșilor săi de oxidare - acizi azotic și azot. Nu există aminoacizi, hidrogenul sulfurat se găsește în cantități mici, există mult oxigen în apă, mineralizarea are loc datorită oxidării complete a materiei organice. Diversitatea speciilor de alge aici este mai mare decât în subzona anterioară, dar numărul și biomasa organismelor sunt mai mici. Cele mai caracteristice acestei subzone sunt diatomeele din genurile Melosira, Diatom, Navicula și verdele din genurile Cosmarium, Spirogyra, Cladophora, Scenedesmus.
În zona oligosaprobică nu există hidrogen sulfurat, există puțin dioxid de carbon, cantitatea de oxigen se apropie de saturația normală și practic nu există substanțe organice dizolvate. Această zonă este caracterizată de o mare diversitate de specii de alge, dar abundența și biomasa lor nu sunt semnificative.
Sistemul lui R. Kolkwitz și M. Marsson a fost îmbunătățit prin extinderea listei și clarificarea tipurilor de indicatori de poluare, precum și prin conversia evaluărilor calitative în cele cantitative (indicele de saprobitate după R. Pantle și G. Bouc). O listă a speciilor de alge - indicatori ai gradului de poluare a corpurilor de apă poate fi găsită în literatura de specialitate (Algae-indicators..., 2000).
unde h este frecvența de apariție a speciei;
s - valoare saprobică.
Valoarea (e) saprobică este exprimată în valori de la 0 la 4 (Pantle și Buck, 1955):
χ (xenosaprobic) - 0;
o (oligosaprobic) - 1;
β (β-μesosaprobitate) - 2;
α (α-μesosaprobitate) - 3;
p (polisaprobicitate) - 4.
Următoarele valori sunt acceptate pentru zonele de tranziție (Sladeček, 1967, 1973):
χ-o (0,4); p-a (2,4);
o-χ (0,6); a-β (2,6);
χ-β (0,8); β-р (2,8);
o-p (1,4); α-р (3,4);
p-o (1,6); p-α (3,6).
Prelucrarea materialului colectat și identificat
Rezultatele obţinute la determinarea algelor sunt prezentate ca o listă sistematică. Principalele cerințe pentru orice sistem de transmitere a informațiilor, inclusiv nomenclatura științifică, sunt universalitatea, unicitatea și stabilitatea. Aceste trei cerințe de bază ale sistemului de comunicare folosit de taxonomiști corespund unui set de reguli – Codul Internațional de Nomenclatură Botanică (ICBN), care a fost adoptat la al VII-lea Congres Internațional de Botanică (Stockholm, 1950). Respectarea regulilor ICBN este obligatorie pentru toți botanicii; încălcarea acestor prevederi poate duce la instabilitatea nomenclaturii botanice.
În taxonomia algelor se disting grupuri taxonomice de organisme (taxa), care sunt acceptate și în taxonomia plantelor superioare. Terminarea numelor tuturor taxonilor de același rang este standardizată după cum urmează:
departament (divizio), -phyta
clasa (classis), -phyceae
ordine (ordo), -ales
familie (familia), -aceae
specii.
Se disting adesea taxoni de rang intraspecific - subspecii, soiuri (varietas), formă (forma) și uneori și subclase (-phycidae), subordine (-ineae) și alte categorii.
Fiecare specie aparține în mod necesar unui gen, un gen unei familii, o familie unui ordin, un ordin unei clase, o clasă unei diviziuni și o diviziune unui regat. Specie așa cum este definită de botanistul rus V.L. Komarova este o colecție de organisme înrudite caracterizate prin caracteristici morfofiziologice și ecologice-geografice specifice acestora. Toți indivizii aceleiași specii se caracterizează printr-o origine filogenetică comună, același tip de metabolism și același habitat.
Specia are un nume format din două cuvinte (principiul nomenclaturii binare). De exemplu: Anabaenaflos- aquae(Lyngb.)Bréb. Primul cuvânt este numele genului, indicând faptul că în natură există un grup de specii înrudite. Al doilea cuvânt, epitetul specific, reflectă caracteristica care distinge o anumită specie de alte specii ale genului. Numele speciei trebuie să fie însoțit de numele de familie al autorului care a descris specia. Numele autorilor sunt scrise prescurtat.
Folosind o listă sistematică de alge, se poate identifica structura fitoplanctonului, diversitatea speciilor de familii și ordinele și diviziunile algelor. În analiza taxonomică, ecologică și geografică a algelor, este necesar să se indice astfel de caracteristici ale speciei precum saprobitatea, habitatul, aciditatea și distribuția geografică. Multe caracteristici sunt indicate în ghidurile de alge atunci când descriem fiecare specie.
Determinarea algelor a fost efectuată la Institutul de Probleme Biologice a Criolitozonei SB RAS folosind determinanți interni și străini.
Capitolul 3. Fitoplanctonul Lacului Soldatskoye
3.1. Compoziția taxonomică a fitoplanctonului
Am găsit 102 specii în planctonul lacului, aparținând la 58 de genuri, 37 de familii, 21 de ordine, 14 clase și 9 diviziuni de alge (Anexa 1). În ceea ce privește numărul de specii, au predominat algele diatomee (41 de specii), verzi (26) și albastru-verzi (14) (Tabelul 2). Streptofitele (6 specii), euglenaceae (5), aurii și galben-verzi (4 specii fiecare) au fost în număr mic. Au fost întâlnite sporadic alge eustigmatofite și dinofite. Speciile de alge albastre-verzi ale genului s-au dezvoltat masiv Oscilatorie, practic priveliștea Oscillatoriaproboscidea.
masa 2
Spectrul taxonom al algelor fitoplanctonului din Lac. Soldatskoye
|
ordine de mărime |
familii |
||||||||||||||
|
Cyanophyta |
|||||||||||||||
|
Euglenophyta |
|||||||||||||||
|
Chrysophyta |
|||||||||||||||
|
Xanthophyta |
|||||||||||||||
|
Eustigmatophyta |
|||||||||||||||
|
Bacillariophyta |
|||||||||||||||
|
Dinophyta |
|||||||||||||||
|
Chlorophyta |
|||||||||||||||
|
Streptophyta |
|||||||||||||||
Lacul este împărțit în 2 secțiuni inegale. Din punct de vedere al diversităţii speciilor, se remarcă situl nr. 1, în care au fost găsite 82 de specii, iar în situl nr. 2 - 63 specii (Tabelul 2). Această distribuție poate fi explicată prin faptul că, conform indicatorilor hidrochimici, situl nr. 1 este mai puțin poluat față de situl nr. 2 (Anexa 2). Prezența nutrienților determină dezvoltarea masivă a anumitor specii și, prin urmare, inhibă dezvoltarea altora. De asemenea, este importantă dimensiunea suprafeței apei și creșterea ei excesivă cu vegetație acvatică mai mare. În lacul nostru, lintia de rață a acoperit aproape complet zona nr. 2 în comparație cu zona nr. 1, reducând astfel fluxul de lumină solară în coloana de apă.
Atunci când sunt determinate, probele prelevate în iunie diferă vizual în numărul de celule de la probele prelevate în iulie. Scăderea numărului de celule este asociată cu dezvoltarea masivă a zooplanctonului, deoarece fitoplanctonul este hrana pentru acestea.
3.2. Speciile – indicatori de saprobitate
Saprobitatea este un complex de proprietăți fiziologice și biochimice ale unui organism care determină capacitatea acestuia de a trăi în apă cu unul sau altul conținut de substanțe organice, adică cu unul sau altul grad de poluare.
Am găsit 59 de specii saprobice în plancton, ceea ce reprezintă 57,8% din numărul total de specii (Anexa 1). Speciile saprobice cu un coeficient de 2 sau mai mult au constituit 44 de specii: β-mezoaprobe - 30 specii, α-β-mezoaprobe - 3 specii, β-α-mezoaprobe - 5 specii, α-mezoaprobe - 6 specii, p-α - 1 specie. Este imposibil de calculat indicele de saprobitate fără indicatori cantitativi, dar din compoziția acestor specii putem spune că indicele de saprobitate va depăși 2, ceea ce clasifică apa ca a treia clasă de puritate cu categoria puțin poluată (Alge-indicatori). ..., 2000).
Această lucrare a fost realizată în combinație cu indicatori hidrochimici și hidrobiologici. În stadiul actual, lacul este ușor transformat; în probele de apă au fost găsite concentrații mari de compuși, indicând acumularea unui număr de compuși biogeni și organici în apă. Acest lucru a condus la dezvoltarea masivă a algelor albastre-verzi, precum și a zooplanctonului.Apele acestui rezervor pot fi folosite pentru uzuri culturale, casnice și piscicole numai cu purificare suplimentară. Pentru a conserva lacul, trebuie efectuate următoarele tipuri de lucrări afară:
- curatarea mecanica a zonei lacului;
- controlează calitatea apei;
- îmbunătățirea zonei de coastă a lacului;
- lucrare de introducere.
Ca urmare a ansamblului de lucrări, se va îmbunătăți alimentația lacului, se va îmbunătăți calitatea apei din lac, iar pe teritoriul adiacent lacului se va crea un mediu sanitar și ecologic favorabil, care va permite lacul pentru a fi menținute în bune condiții sanitare în viitor.
pentru a reface lacul. Soldatsky
|
EVENIMENT |
IMPLEMENTAREA |
TERMEN |
INTERPRETURI |
|
Curățarea mecanică a zonei lacului |
Curățarea zonelor de coastă și de apă de deșeurile menajere. |
perioada de dezvoltare intensivă a plantelor |
voluntari |
|
Controlul calității apei |
Prelevarea probelor de apă |
perioada de apă deschisă (mai-septembrie 2017-2020) |
Gerasimenko S., |
|
Îmbunătățirea zonei de coastă a lacului |
1. Nivelarea și întărirea versanților de mal a lacului prin însămânțarea de iarbă peste un strat de pământ vegetal folosind o geogrilă. |
iunie-august 2019-2020 |
districtul Guba, |
|
Lucrare de introducere |
Introducerea de obiecte biologice pentru a îmbunătăți starea apei lacului |
perioada de apă deschisă |
administrația școlii nr. 21, |
|
Lucrări de propagandă pentru păstrarea stării ecologice a lacurilor |
Discurs la conferințe la nivel de școală, oraș și republican |
octombrie-ianuarie 2017-2020 |
elevii școlii nr.21 |
CONCLUZIE
Fitoplanctonul lacului Soldatskoye este reprezentat de 102 specii aparținând a 58 de genuri, 37 de familii, 21 de ordine, 14 clase și 9 divizii de alge. Speciile de alge albastre-verzi ale genului s-au dezvoltat masiv Oscilatorie, practic priveliștea Oscillatoriaproboscidea. Situl nr. 1 este mai bogat în diversitate de specii comparativ cu situl nr. 2. După tipurile de indicatori, apa lacului aparține clasei a treia de puritate.
Pentru a conserva lacul, este necesar să se efectueze curățarea mecanică a rezervorului; îmbunătățirea zonei de coastă și efectuarea lucrărilor de introducere.
Pentru a crea condiții favorabile pe acest rezervor, este necesar să se efectueze lucrări de restaurare cu implicarea publicului, a serviciilor de locuințe și comunale și a voluntarilor reprezentați de elevii școlii nr. 21 și locuitorii districtului Gubinsky. Intenționăm să continuăm cercetările în viitor. Sperăm din suflet că, prin eforturi comune, vom crea un loc frumos în care locuitorii să se relaxeze.
Bibliografie
Indicatori de alge în evaluarea calității mediului. Partea I. Barinova S.S. Aspecte metodologice ale analizei diversităţii biologice a algelor. Partea a II-a. Barinova S.S., Medvedeva L.A., Anisimova O.V. Caracteristicile ecologice și geografice ale algelor indicator. - Moscova: VNIIprirody, 2000. - 150 p.
Alge: Director / Vasser S.P., Kondratyeva N.V., Masyuk N.P. și altele - Kiev: Nauk. Dumka, 1989. - 608 p.
Ermolaev V.I. Fitoplanctonul rezervoarelor din bazinul lacului Sartlan. - Novosibirsk: Știință, 1989. - 96 p.
Ivanova A.P. Algele lacurilor urbane și suburbane ale Văii Lenei Mijlocii. - rezumatul autorului disertație pentru concursul academic Etapa. Ph.D. biol. Sci. - Moscova, 2000. - 24 p.
Kuzmin G.V. Algele plancton din Sheksninsky și părțile adiacente ale rezervorului Rybinsk // Biologia, morfologia și sistematica organismelor acvatice. - Moscova: Știință, 1976. - Numărul. 31 (34). - P. 3-60
Kuzmin G.V. Tabele pentru calcularea biomasei de alge. Pretipărire. - Magadan, 1984. - 48 p.
Lasukov R.Yu. Locuitorii lacurilor de acumulare. Identificator de buzunar. - Moscova: Țara Pădurii, Editura. 2, rev., 2009. - 128 p.
Makarova I.V., Pichkily A.O. Pe unele aspecte ale metodologiei de calcul a biomasei fitoplanctonului // Botan. j-l. - 1970. - T. 55, nr. 10. - P. 1488-1494.
Pantle F., Buck H. Die biologischeüberwachung der Gewasser und die Darstellung der Ergebnisse. Gaz.- und Wasserbach. - 1955. - Bd.96, Nr. 18. - S. 1-604.
SladečekV. 1973.Sistemul calităţii apei din punct de vedere.Ergebn.limnol. - 7: 1-128.
Anexa 1
Lista sistematică a algelor din Lacul Soldatskoye
|
Alge |
Sectiunea 1 |
Sectiunea 2 |
Habitat |
Ga-frunte |
Sa-probabilitate |
|
CIANOFITĂ |
|||||
|
ClasăCyanophyceae |
|||||
|
OrdinSinecococi |
|||||
|
Familie Merismopediaceae |
|||||
|
Merismopediaglauca(Ehr.) Näg. |
|||||
|
Merismopediamajor(Smith) Geitl. |
|||||
|
OrdinChroococcales |
|||||
|
FamilieMicrocystaceae |
|||||
|
Microcystisaeruginosa Kütz. amenda. Elenk. |
|||||
|
Microcystispulverea f. planctonica(G. W. Smith) Elenk. |
|||||
|
FamilieAphanothecaceae |
|||||
|
Aphanothecesaxicola Ponei. |
|||||
|
OrdinOscillatoriales |
|||||
|
FamilieOscillatoriaceae |
|||||
|
Oscillatoria acutissima Kuff. |
|||||
|
Oscillatoriaamphibia Ag. f. amfibii |
|||||
|
Oscillatoriachalybea(Mert.) Gom. |
|||||
|
Oscilatorielimoza Ag. |
|||||
|
Oscilatorieplanctonica Wolosz. mult |
|||||
|
Oscillatoriaproboscidea Gom. mult |
|||||
|
Oscillatoriapseudogeminata G. Schmid |
|||||
|
OrdinNostocales |
|||||
|
Familie Aphanizomenonaceae |
|||||
|
Aphanizomenonflos-aquae(L.) Ralphs |
|||||
|
FamilieNostocaceae |
|||||
|
Anabaena flos-aquae(Lyngb.) Breb. |
|||||
|
EUGLENOPHYTA |
|||||
|
ClasăEuglenophyceae |
|||||
|
Ordin Euglenales |
|||||
|
FamilieEuglenaceae |
|||||
|
Trachelomonashispida(Perty) Stein amend. Defl. |
|||||
|
Euglena granulata var. polimorfe(Dang.) Popova |
|||||
|
Euglena hemichromatata Skuja |
|||||
|
Euglena viridis Ehr. |
|||||
|
Phacusstriatus Franţa |
|||||
continuarea anexei 1
|
HRISOFITA |
|||||
|
ClasăChrysophyceae |
|||||
|
OrdinCromulinale |
|||||
|
FamilieDinobryonaceae |
|||||
|
Dinobryonsociale Ehr. |
|||||
|
ClasăSynurophyceae |
|||||
|
OrdinSynurales |
|||||
|
FamilieSynuraceae |
|||||
|
Mallomonas denticulat Matv. |
|||||
|
Mallomonas longiseta Lemm. |
|||||
|
Mallomonasradiata Conrad |
|||||
|
XANTOFITA |
|||||
|
ClasăXanthophyceae |
|||||
|
OrdinMischococcales |
|||||
|
FamilieBotrydiopsidaceae |
|||||
|
Botrydiopsiseriensis Zăpadă |
|||||
|
FamiliePleurochloridaceae |
|||||
|
Chloridella neglecta(Pasch. et Geitl.) |
|||||
|
Nephrodiella lunaris Paştele. |
|||||
|
OrdinTribonematales |
|||||
|
FamilieTribonemataceae |
|||||
|
Tribonemaaequale Paştele. |
|||||
|
EUSTIGMATOPHYTA |
|||||
|
ClasăEustigmatophyceae |
|||||
|
OrdinEustigmatales |
|||||
|
FamiliePseudocharaciopsidaceae |
|||||
|
Elipsoidionregulare Paştele. |
|||||
|
BACILARIOFITĂ |
|||||
|
Clasă Coscinodiscophyceae |
|||||
|
OrdinAulacoseirales |
|||||
|
FamilieAulocosiraceae |
|||||
|
Aulocosiraitalica(Kütz.) Simon. |
|||||
|
Clasă Mediophyceae |
|||||
|
OrdinThalassiosirales |
|||||
|
FamilieStephanodiscaceae |
|||||
|
Cyclotella meneghiniana Kütz. |
|||||
|
Cyclotella sp. |
|||||
|
Handmanniacomta(Ehrenb.) Kociolek și Khursevici |
|||||
|
Clasă Bacillariophyceae |
|||||
|
OrdinAraphales |
|||||
|
FamilieFragilariaceae |
|||||
|
Asterionellaformosa Hassall |
|||||
|
Fragilaria capucina Desm. |
|||||
|
Fragilariaintermedia Grun. |
|||||
|
Ulnaria ulna(Nitzsch) Compère |
|||||
|
FamilieDiatomaceae |
|||||
continuarea anexei 1
|
Diatom vulgar Bory |
|||||
|
FamilieTabellariaceae |
|||||
|
Tabellaria fenestrata(Lyngb.)Kütz. |
|||||
|
OrdinRaphales |
|||||
|
FamilieNaviculaceae |
|||||
|
Caloneissilicula(Ehr.) Cl. |
|||||
|
Hipodontacapitata(Ehrenb.) Lange-Bert., Metzeltin et Witkowski |
|||||
|
Naviculacryptocephala Kütz. |
|||||
|
Navicula cuspidate f. primigena Dipp. |
|||||
|
Naviculadigitoradiata(Greg.) A.S. |
|||||
|
Naviculaelginensisvar.cuneata(M. Möller) Lange-Bertalot |
|||||
|
Naviculamutica Kütz. |
|||||
|
Naviculaoblonga Kütz. |
|||||
|
Navicularradiosa Kütz. |
|||||
|
Pinnulariagibba var. linearis Hust. |
|||||
|
Pinnulariaviridis var. eliptica Meist. |
|||||
|
Sellaphoraparapupula Lange-Bert. |
|||||
|
Stauroneisphoenicenteron Ehr. |
|||||
|
FamilieAchnanthaceae |
|||||
|
Achnanthesconspicua A. Mayer |
|||||
|
Achnantheslanceolata var. eliptica Cl. |
|||||
|
Achnantheslinearis(W. Sm.) Grun. |
|||||
|
Cocconeisplacentula Ehr. |
|||||
|
Planothidium lanceolatum(Bréb. ex Kütz.) Lange-Bert. |
|||||
|
FamilieEunotiaceae |
|||||
|
Eunotiafaba(Ehr.) Grun. |
|||||
|
FamilieCymbellacee |
|||||
|
Amfora ovalis Kütz. |
|||||
|
Cymbellacymbiformis(Ag. ?Kütz.) V.H. |
|||||
|
Cymbellaneocistula Krammer |
|||||
|
Cymbellumida(Bréb.) V.H. |
|||||
|
FamilieGomphonemataceae |
|||||
|
Gomphonemaacuminatum var. coronatum(Ehr.) W. Sm. |
|||||
|
Gomphonemacapitatum Ehrenb. |
|||||
|
Gomphonemahelveticum Brun. |
|||||
|
Gomphonemaparvulum(Kütz.) Grun. |
|||||
|
FamilieEpithemiaceae |
|||||
|
Epithemiaadnata(Kütz.) Breb. |
|||||
|
FamilieNitzschiaceae |
|||||
|
Nitzschiaacicularis W.Sm. |
|||||
|
Nitzschiapalea(Kütz.) W. Sm. |
|||||
|
NitzschiapaleaceaeGrun. |
continuarea anexei 1
|
DINOPHYTA |
|||||
|
ClasăDinophyceae |
|||||
|
Ordin Gonyaulacales |
|||||
|
Familie Ceratiaceae |
|||||
|
Ceratiumhirundinella T. furcoides(Lev.) Schröder |
|||||
|
CHLOROPHYTA |
|||||
|
ClasăChlorophyceae |
|||||
|
Ordin Chlamydomonadales |
|||||
|
FamilieChlamydomonadaceae |
|||||
|
Chlamydomonas sp. |
|||||
|
OrdinSphaeropleales |
|||||
|
FamilieSphaerocystidaceae |
|||||
|
Sphaerocystis planctonica(Korsch.) |
|||||
|
FamilieHidrodictyaceae |
|||||
|
Pediastrumborianum(Turp.) Menegh. |
|||||
|
Pediastrum duplex Meyen var. duplex |
|||||
|
Pediastrum tetra(Ehr.) Ralfs |
|||||
|
Tetraеdroncaudatum(Corda) Hansg. |
|||||
|
Tetraedron minim(A. Br.) Hansg. |
|||||
|
FamilieSelenastraceae |
|||||
|
Monoraphidium contortum(joi) Kom.-Legn. |
|||||
|
Monorafidiu neregulat(G. M. Smith) Kom.-Legn. |
|||||
|
Monoraphidiumkomarkovae Nyg. |
|||||
|
Monoraphidiumminutum(Näg.) Kom.-Legn. |
|||||
|
Mesastrum gracile(Reinsch) T.S. Garcia |
|||||
|
FamilieScenedesmaceae |
|||||
|
Coelastrummicroporum Ponei. |
|||||
|
Crucigeniafenestrata(Schm.) Schm. |
|||||
|
Scenedesmusacuminatus(Lagerh.) Chod. |
|||||
|
Scenedesmusarcuatus(Lemm.) Lemm. |
|||||
|
Scenedesmusellipticus Corda |
|||||
|
Scenedesmusfalcatus Chod. |
|||||
|
Scenedesmusobliquus(Turp.) Kütz. |
|||||
|
Scenedesmusquadricauda(Turp.) Breb. |
|||||
|
Tetrastrumtriunghiular(Chod.) Kom. |
|||||
|
Clasă Oedogoniophyceae |
|||||
|
OrdinOedogoniales |
|||||
|
FamilieOedogoniaceae |
|||||
|
Oedogonium sp. |
|||||
sfarsitul aplicarii 1
|
Clasă Trebouxiophyceae |
|||||
|
Ordin Clorellale |
|||||
|
Familie Chlorellaceae |
|||||
|
Actinastrumhantzschii Lagerh. |
|||||
|
Dictyosphaerium pulchellum Lemn. |
|||||
|
FamilieOocystaceae |
|||||
|
Oocystisborgei Zăpadă |
|||||
|
Oocystis lacustris Chod. |
|||||
|
STREPTOFITA |
|||||
|
K fată Zygnematophyceae |
|||||
|
OrdinZygnematales |
|||||
|
Familie Mougeotiaceae |
|||||
|
Mougeotia sp. |
|||||
|
OrdinDesmidiales |
|||||
|
FamilieClosteriaceae |
|||||
|
Closterium moniliferum(Bory) Ehr. |
|||||
|
FamilieDesmidiaceae |
|||||
|
Staurastrumtetracerum Ralfs |
|||||
|
Cosmarium botrytis Menegh. |
|||||
|
Cosmariumformosulum Hoff |
|||||
|
Cosmarium sp. |
|||||
Notă: habitat: p - plancton, b - bentos, o - încrustare; halobism: i - indiferent, gl - halofil, gb - halophobe, mzb - mezohalob.
Anexa 2
Caracteristicile calității apei de suprafață la locul nr. 1 „SKVER”
Caracteristici ale calității apei de suprafață la situl nr. 2 „PANDA”
- „Cronicile lui Amber”. Cărți în ordine. Recenzii. Roger Zelazny „Cronicile lui Amber Roger Zelazny Cei nouă prinți ai chihlimbarului a continuat
- Ciupercă de orez: beneficii și daune
- Energia umană: cum să vă aflați potențialul energetic Energia vitală umană după data nașterii
- Semne zodiacale pe elemente - Horoscop