
Functions of invagination of the cytoplasmic membrane. Cytoplasmic membrane, its functions and structure
Cell – the basic structural, functional, genetic unit of the organization of living things, an elementary living system. A cell can exist as a separate organism (bacteria, protozoa), or as part of the tissues of multicellular organisms. The term "cell" was proposed by the English explorer Robert Hooke in 1665.
The main provisions of the cell theory were formulated by the botanist Schleiden (1838) and the zoologist-physiologist Schwann (1839). In 1858, Virchow supplemented the provisions with a statement about cell division.
Basic provisions of modern cell theory:
All living organisms are made up of cells. A cell is a unit of structure, functioning, reproduction and individual development of living organisms. There is no life outside the cell.
The cells of all organisms are similar to each other in structure and chemical composition.
Cells can only be formed from cells by division.
The cellular structure of all living organisms is evidence of a unity of origin.
Modern definition of a cell:
a cell is an open biological system, bounded by a semi-permeable membrane, consisting of a nucleus and cytoplasm, capable of self-regulation and self-reproduction.
Two types of organisms have a cellular structure - prokaryotes (bacteria and blue-green algae) and eukaryotes (Fig. 2.1). Eukaryotic cells consist of a surface apparatus (cytoplasmic membrane), cytoplasm and nucleus.
Cytoplasmic membranes
Cytoplasmic membranes perform a number of important functions: barrier (delimiting), regulatory (regulate metabolic flows), transport (ensuring selective permeability of substances through passive and active transport), structural, metabolic. Biological membranes are composed mainly of lipids, proteins and
carbohydrates (Fig. 2.2). Several models of the structure of cytoplasmic membranes have been proposed (the “sandwich” model - the Danieli and Dawson model, the Lenard model, etc.). There appear to be several types of membranes depending on function. Currently, the fluid-mosaic model proposed by Singer-Nicholson (1972) is adopted as a basis. According to this model, the membranes include a bimolecular layer of lipids, which includes protein molecules.
Lipids are water-insoluble substances. They have a polar (charged) head and long uncharged (non-polar) carbohydrate chains. Lipid molecules face each other with non-polar ends, and their polar poles (heads) remain outside, forming hydrophilic surfaces (Fig. 2.3).
Membrane proteins can be divided into three groups: peripheral (most weakly associated with the membrane), submerged (semi-integral) and permeating (integral), forming membrane pores. Functionally, membrane proteins are divided into enzymatic, transport, structural and regulatory.
On the outer surface of the plasma membrane, protein and lipid molecules are associated with carbohydrate chains to form a glycocalyx. Carbohydrate chains act as receptors; the cell acquires the ability to specifically respond to external influences. Thus, the interaction of a hormone with “its” receptor from the outside causes a change in the structure of the integral protein, which leads to the triggering of a cellular response. In particular, such a response can manifest itself in the formation of “channels” through which solutions of certain substances begin to enter or exit the cell.
One of the important functions of the membrane is to ensure contacts between cells within organs and tissues.
Under the plasma membrane on the cytoplasmic side there is a cortical layer and intracellular fibrillar structures that provide the mechanical stability of the membrane.
In plant cells, outside the membrane there is a dense structure - the cell membrane, consisting of polysaccharides (cellulose).
One of the most important properties of the cytoplasm is associated with the ability to pass various substances into or out of the cell. This is necessary to maintain the consistency of its composition. Small molecules and ions pass through membranes by passive and active transport.
Passive transport occurs without energy expenditure through diffusion, osmosis and facilitated diffusion (Fig. 2.4). Diffusion –

transport of molecules and ions through the membrane from an area with high to an area with low concentration, i.e. along the concentration gradient. If substances are highly soluble in fats, then they penetrate the cell by simple diffusion (oxygen, carbon dioxide). The diffusion of water through semi-permeable membranes is called osmosis. Water is also able to pass through membrane pores formed by proteins and transport molecules of ions and substances dissolved in it. Facilitated diffusion is the transport of substances that are insoluble in fats and do not pass through pores through ion channels using carrier proteins.
Active transport of substances across the membrane occurs with the expenditure of ATP energy and with the participation of carrier proteins. It is carried out against a concentration gradient (this is how amino acids, sugar, potassium, sodium, calcium ions, etc. are transported). An example of active transport is the operation of the potassium-sodium pump. The concentration of K inside the cell is 10-20 times higher than outside, and Na - vice versa. To maintain this concentration, three Na ions are transferred from the cell for every two K ions into the cell. This process involves a protein in the membrane, which acts as an enzyme that breaks down ATP, releasing the energy necessary for the pump to operate (Fig. 2.5. A).
The transfer of macromolecules and large particles into the cell is carried out by endocytosis, and removal from the cell by exocytosis.
During endocytosis (Fig. 2.5. B), the membrane forms invaginations or outgrowths, which then lace up and transform into intracellular vesicles containing the product captured by the cell. This process occurs with the expenditure of ATP energy. There are two types of endocytosis - phagocytosis (absorption of large particles by the cell) and pinocytosis (absorption of liquid substances).
The membrane takes part in the removal of substances from the cell during the process of exocytosis. In this way, hormones, proteins, fat droplets, etc. are removed from the cell.
Beneath the bacterial cell wall is the cytoplasmic membrane (CPM). It separates the cell contents from the cell wall and is an essential structure of any cell.
The thickness of the bacterial CPM is usually about 6-8 nm. It accounts for up to 15% of the dry mass of the cell. It consists of lipids (15-45%), proteins (45-60%) and a small amount of carbohydrates (about 10%). Lipids are represented by phospholipids - up to 30% of the dry weight of the membrane. Among them, phosphatidyl glycerol and diphosphatidyl glyceride (cardiolipin) predominate - an essential component of the mitochondrial membranes of eukaryotes. Smaller quantities contain phosphatidylinositol and phosphatide yl-
ethanolamine In addition to phospholipids, various glycolipids, small amounts of carotenoids and quinones were found in the membrane. In the composition of lipids derived from glycerol, fatty acids atypical for membranes were identified - saturated or monounsaturated with 16-18 carbon atoms, as well as acids not found in eukaryotic membranes - cyclopropane and branched fatty acids with 15-17 carbon atoms. The set of fatty acids, as well as the membrane lipids consisting of them, is species specific for prokaryotes.
Membrane lipids are small polar molecules bearing hydrophilic (heads) and hydrophobic (tails) groups. In an aqueous environment, they spontaneously form a closed bimolecular layer - a bilayer. This layer serves as a significant barrier to ions and polar compounds. Organized into a bimolecular layer, lipids form the structural basis of the membrane, maintain mechanical stability and impart hydrophobicity to it.
Proteins make up more than half of the dry mass of the membrane. There are more than 20 different types. Based on differences in the strength of bonds with lipids and location in the membrane, proteins are divided into integral and peripheral. Integral proteins are immersed in the hydrophobic region of the membrane, where they form numerous bonds with the hydrocarbon chains of lipids,
creating lipoprotein complexes. Peripheral proteins are localized on the surface of the hydrophilic layer and are often attached to integral proteins (Fig. 3.14).
Fig.3.14. Structure of the cytoplasmic membrane: 1 - lipids; 2 - glycoproteins; 3 - peripheral proteins; 4 - integral proteins
Based on their functions within membranes, membrane proteins can be divided into two groups: structural and dynamic.
The functions of structural proteins are limited to maintaining the structural integrity of the membrane. They are located on the surface of the hydrophilic lipid layer, acting as a molecular bandage.
Dynamic proteins include proteins that are directly involved in all processes occurring on the membrane. They are divided into three classes: transport, involved in the transport of compounds in and out of the cell; catalytic, performing the functions of enzymes in reactions occurring on the membrane; receptor proteins that specifically bind certain compounds (toxins, hormones) on the outside of the membrane.
Carbohydrates in the membrane are not in a free state, but are interconnected with proteins and lipids into glycoproteins. They're like
As a rule, they are localized only on the outer surface of the membrane and function as recognition receptors for environmental factors.
The cytoplasmic membrane of bacteria, like all other biological membranes, is an asymmetric liquid crystalline structure. The asymmetry is due to differences in the chemical structure of protein molecules and their location in the lipid bilayer of the membrane. Some proteins are located on the surface of the bilayer, others are immersed in its thickness, and others pass through from the inner to the outer surface of the bilayer. The strictly defined orientation of membrane proteins, in turn, is due to the fact that they are synthesized and incorporated into the membrane asymmetrically. The outer and inner surfaces of the membrane also differ in enzymatic activity. Depending on conditions (for example, temperature), the CPM can be in different phase states: liquefied or crystalline. During the transition of one liquid crystalline phase to another, the mobility of the membrane components and its packing density change, which, in turn, leads to a disruption of its functional activity.
Structural organization and functions of the cytoplasmic membrane. To explain the nature and mechanism of the numerous functions of the CPM, the most suitable is the fluid-mosaic model of the organization of biological membranes, proposed by R. Singer and A. Nicholson in 1972. According to this model, membranes are two-dimensional solutions of globular proteins and lipids oriented in a certain way. Lipids form a bilayer in which the hydrophilic “heads” of the molecules face outward, and the hydrophobic “tails” are immersed in the thickness of the membrane, while possessing sufficient flexibility. Membrane lipids and many proteins move freely in the bilayer, but only in the lateral direction (lateral diffusion). In the transverse direction, i.e., from one surface of the membrane to the opposite, proteins cannot move, and lipids move extremely slowly (once every few hours). The reason for the absence or low activity of transverse diffusion appears to be the asymmetric distribution of lipids:
Some lipids are more abundant in the outer part of the bilayer, while others are more abundant in the inner part. The consequence of this is the unequal electron density (conductivity) of the bilayer in the transverse direction.
The CPM is in a liquid crystalline or liquefied state only under certain, so-called
biological temperatures. When the temperature decreases (below the melting point, Tm), the lipids transform into a crystalline state, the degree of viscosity increases until the membrane hardens. The temperature that causes the membrane to harden is determined by the content of unsaturated and
branched fatty acids. The more of them there are in the membrane, the lower the temperature of the transition of lipids from the liquid crystalline state to the crystalline state.
Prokaryotes have the ability to regulate membrane fluidity by changing the number of double bonds and the chain length of fatty acid molecules. Thus, in E. coli, when the environmental temperature decreases from 42° C to 27° C, the ratio of saturated and unsaturated fatty acids in the membrane decreases from 1.6 to 1.0, i.e., the content of unsaturated fatty acids reaches the level of saturated ones. This prevents an increase in viscosity and ensures that cells maintain physiological activity at low temperatures.
The CPM performs numerous vital functions in prokaryotes. They are mainly determined by proteins localized in it, which act as channels, receptors, energy regenerators, enzymes, transport functions and others. The CPM is the main osmotic barrier, which, due to the presence of membrane transport mechanisms, selectively transports substances into the cell and removes metabolic products from it. The selective permeability of the CPM is due to substrate-specific permeases localized in it, which actively transport various organic and mineral substances through the membrane. The CPM contains enzymes for the biosynthesis of membrane lipids and macromolecules that make up the cell wall, outer membrane, and capsule. The CPM is the site of localization of redox enzymes that carry out
electron transport, oxidative and photosynthetic phosphorylation, electrochemical energy generation
transmembrane potential (A// +) and chemical (ATP). CPM
n
performs important functions in the biosynthesis and translocation of secreted proteins by gram-negative bacteria. The biosynthesis of these proteins is carried out on ribosomes attached to the CPM. Gram-negative bacteria have special receptor proteins on the CPM that “recognize” signals from the large ribosomal subunit about ribosome attachment and the beginning of protein synthesis. Membrane receptor proteins interact with the large subunit of the ribosome, forming a ribosome-membrane complex on which the synthesis of secreted proteins is carried out. In this way, for example, E. coli synthesizes alkaline phosphatase, Bac. subtilis - a-amylase. The CPM also ensures the transfer of these proteins into the periplasmic space. The CPM plays a great role in the regulation of cell division, replication of chromosomes and plasmids, and the subsequent segregation of these genetic elements between newly formed daughter cells.
All prokaryotes, along with the cytoplasmic membrane, contain its derivatives - intracellular membranes that perform specialized functions. The cytoplasmic membrane is capable of forming all kinds of invaginations (invaginations). These invaginations constitute intracellular membranes, which have different lengths, packaging, and localization in the cytoplasm. They can be collected into complex balls - lamellar, honeycomb or tubular formations. Less complex membranes take the form of simple loops or tubules of varying lengths. Regardless of the complexity of the organization of intracellular membranes, they are all derivatives of the cytoplasmic membrane. The size of their active surface exceeds that of the cytoplasmic membrane. This gives grounds to judge the great functional activity of these structures in cells.
A particularly rich intracellular membrane apparatus is found in nitrogen-fixing and photosynthetic bacteria, Brucella, and nitrifying bacteria. Photosynthetic bacteria (Rhodospirillum rubrum) have membranes that look like closed bubbles - vesicles. Their formation begins with invagination of the cytoplasmic membrane, which then forms a tube. Constrictions appear on the tube, dividing it into a series of bubbles. These vesicles are called chromatophores. They contain light-absorbing pigments - bacteriochlorophylls and carotenoids, electron transport enzymes - ubiquinones and cytochromes, components of the phosphorylation system. In some photosynthetic prokaryotes, in particular in purple sulfur bacteria and cyanobacteria, the photosynthetic apparatus is represented by stacks of membranes that have a flattened shape and, by analogy with the grana of chloroplasts of green plants, are called thylakoids (Fig. 3.15).  They concentrate photosynthetic pigments, enzymes of the electron transport chain and phosphorylation systems. A feature of cyanobacterial thylakoids is the lack of connection with the cytoplasmic membrane. This is the only group of prokaryotes that has a differentiated membrane system.
They concentrate photosynthetic pigments, enzymes of the electron transport chain and phosphorylation systems. A feature of cyanobacterial thylakoids is the lack of connection with the cytoplasmic membrane. This is the only group of prokaryotes that has a differentiated membrane system.
In nitrifying bacteria, the intracellular membrane apparatus has the form of plates, or lamellae, consisting of flat vesicles (Fig. 3.16).  Of the intracellular membranes, mesosomes have the most complex structure. They are spirally twisted, flat or spherical tubular bodies. Mesosomes are formed during cell division in the zone of formation of the transverse septum. They take part in chromosome replication and distribution of genomes between daughter cells, and in the synthesis of cell wall substances. To participate
Of the intracellular membranes, mesosomes have the most complex structure. They are spirally twisted, flat or spherical tubular bodies. Mesosomes are formed during cell division in the zone of formation of the transverse septum. They take part in chromosome replication and distribution of genomes between daughter cells, and in the synthesis of cell wall substances. To participate
mesosomes in cell division are indicated by its connection with the DNA of the nucleoid. Well-developed mesosomes are found only in gram-positive bacteria.
The information accumulated to date suggests that the membrane structures of bacteria are sufficiently differentiated and ensure the course of various metabolic processes in the cell.
- Cytoplasm and cytoplasmic inclusions
In the cytoplasm, two fractions are distinguished. One of them presents the structural elements: ribosomes, aerosomes,
carboxysomes, storage inclusions, genetic apparatus, Another fraction contains a complex mixture of soluble RNA, enzyme proteins, pigments, minerals, products and substrates of metabolic reactions. This fraction is called cytosol.
Due to the presence of various organic compounds, the cytoplasm of bacterial cells is characterized by increased viscosity. It is 800-8000 times greater than the viscosity of water (approaching the viscosity of glycerin). Young cells that are in lag phase or in the early stages of logarithmic phase have a lower cytoplasmic viscosity; in aging people, the viscosity increases, resembling a gel in consistency. The degree of cytoplasmic viscosity characterizes not only the age of the cell, but also its physiological activity. An increase in cytoplasmic viscosity in old cultures is one of the factors responsible for a decrease in the physiological activity of cells. Cytoplasm is the medium that connects all intracellular structures into a single system.
Ribosomes. The cytoplasm of a bacterial cell constantly contains spherical structures, 15-20 nm in size, with a molecular weight of 3106.
Ribosomes consist of 60-65% ribosomal RNA and 35-40% protein. The latter are rich in basic amino acids. During ultracentrifugation, bacterial ribosomes settle at a rate of about 70 Svedberg units (S)7, which is why they are called 708-ribosomes. Cytoplasmic ribosomes of eukaryotes are larger and are called 80S ribosomes (their sedimentation constant is 80S).
Each ribosome consists of two subunits: 30S and 50S, which differ in the size of the RNA molecules and the amount of protein they contain. The large subunit (50S) contains two rRNA molecules - 5S and 23S and 35 molecules of various proteins. The small subunit (30S) includes one molecule of 16 rRNA and 21 molecules of different types of proteins. The number of ribosomes in a cell is not constant - from 5,000 to 90,000. It is determined by the age of the cell and the conditions of bacterial cultivation. The minimum amount is contained at the beginning of the lag phase, and the maximum - in the exponential phase of culture growth. In E. coli, during the period of active growth on a complete nutrient medium, 5-6 ribosomes are synthesized in 1 second. Most of them in the cytoplasm of bacteria are in a free state, and the rest is
S = 1 swedberg unit = 10"13 cm (s) field unit.
united by strands of messenger RNA into polysomes. The number of ribosomes in polysomes can reach several dozen. This indicates a high protein-synthesizing activity of the cell, since ribosomes are the site of protein synthesis. They are figuratively called protein “factories”.
Gas vacuoles (aerosomes). These structures are characteristic only of some water and soil bacteria. They are found in phototrophic sulfur bacteria, colorless filamentous bacteria, as well as in bacteria of the genus Renobacter. There are up to 40-60 of them in a cell (Fig. 3.17). Gas vacuoles are surrounded by thin 
Rice. 3.17. Renobacter vocuolatum cell with aerosomes (magnification x 70,000)
protein membrane. They contain gas bubbles, the number of which is not constant. The composition and pressure of gas in bubbles and aerosomes are generally determined by the amount of gases dissolved in the environment. Aerosomes are either in a compressed state or filled with a gas medium. Their condition is regulated by hydrostatic pressure of the environment. A sharp increase in pressure causes compression of aerosomes and the cells lose their buoyancy.
Aerosomes regulate the buoyancy of the cell, providing the ability to move it to favorable conditions of aeration, lighting, and nutrient content. A special feature is their one-time operation when filled with gas. After compression under the influence of hydrostatic pressure, they are not refilled with gas and
are gradually destroyed. The cell can reproduce them only by forming them anew.
When the aerosomes are filled with gas, the bacteria are held on the surface of the water; when they are compressed, they sink into its thickness or settle to the bottom of the reservoir. This unique method of movement was developed in the process of evolution mainly in bacteria that lack flagella and, consequently, the ability to actively move.
Phycobilisomes. These intracellular structures are characteristic of cyanobacteria. They have the form of granules with a diameter of 28-55 nm and are the site of localization of water-soluble pigments - phycobiliproteins, which determine the color of cyanobacteria and are involved in photosynthesis.
Chlorosomes, or chlorobium vesicles, are structures in which the photosynthetic apparatus of green bacteria of the genus Chlorobium is localized. They have an elongated shape, 100-150 nm long, 50-70 nm wide, surrounded by a single-layer protein membrane. Chlorosomes are located in a dense layer under the cytoplasmic membrane, but are physically separated from it. The chlorosomes of green bacteria contain photosynthetic pigments - bacteriochlorophylls, which absorb light quanta and transfer energy to the reaction centers of photosynthesis.
Carboxysomes. The cells of certain types of phototrophic (cyanobacteria, some purple bacteria) and chemolithotrophic (nitrifying bacteria) prokaryotes contain polyhedron-shaped structures with a size of 90-500 nm. In accordance with the function they perform, they are called carboxysomes. They contain the enzyme ribulose diphosphate carboxylase, which catalyzes the binding reaction of carbon dioxide with ribulose diphosphate in the Calvin cycle. In autotrophic bacteria they are the site of carbon dioxide fixation. Carboxysomes are surrounded by a single-layer protein membrane, which protects the enzyme from the effects of intracellular proteases.
Spare nutrients* In addition to the described structural elements, the cytoplasm of bacteria contains granules of various shapes and sizes in the form of inclusions. Their presence in
cell is not constant and is associated with the composition of the nutrient medium and the physiological state of the culture. Many cytoplasmic inclusions consist of compounds that serve as a source of energy and a source of nutrients. They are usually formed in cultures on fresh, nutrient-rich media, when cell growth is inhibited for some reason, or after the end of a period of active growth. The chemical composition of inclusions is different and not the same in different types of bacteria. They can be polysaccharides, lipids, crystals and granules of inorganic substances.
Of the polysaccharides, we should first of all mention starch, glycogen and a starch-like substance - granulosa. The most common is glycogen. It is found in bacilli, salmonella, Escherichia coli, sardines, etc. In spore-bearing anaerobes of the genus Clostridium, the cells contain small granulosa granules. These inclusions are used by the cell as sources of energy and carbon.
Lipids accumulate in the cytoplasm of bacteria in the form of small drops and grains. In many bacteria, lipid inclusions are represented by poly-p-hydroxybutyric acid, which often accounts for up to 50% of the dry biomass of bacteria. Bacteria of the genus Bacillus and phototrophic bacteria are especially rich in this compound. Poly-p-hydroxybutyric acid is synthesized in large quantities during the growth of microorganisms on media rich in carbohydrates. In each polylactide chain, p-hydroxybutyric acid residues account for up to 60%, and therefore this compound is an ideal “storehouse” of energy for bacteria. Some microorganisms accumulate waxes and neutral fats (triglycerides). Thus, in mycobacteria and actinomycetes, waxes sometimes make up up to 40% of the dry mass; yeast cells of the genus Candida and Rhodotorula are rich in neutral fats; their number reaches almost 60%.
All lipid inclusions in microorganisms serve as a source of energy and carbon.
In the cells of many bacteria, special inclusions called volutin grains are often found. By chemical nature, volutin is a polyphosphate. Name volutin
comes from the species name of the sulfur bacteria Spirillum volutans, in which these inclusions were first described. Volutin has the property of metachromasia, i.e. causes color changes in some dyes. If bacteria are stained with methylene blue or toluidine blue, the volutin grains become purple or red-violet in color. In this regard, researchers V. Babes and E. Ernst, who first described these inclusions, called them metachromatic grains. Volutin grains have a spherical shape, up to 0.5 microns in size. They are formed under conditions of good nutrition of microorganisms, especially in media rich in carbohydrates, as well as in the presence of glycerol in the environment. Volutin is found in the cells of both pathogenic and saprophytic bacteria, for example, spirilla, azotobacter, and the causative agent of diphtheria.
Volutin is used by the cell mainly as a source of phosphate groups and partly energy.
In colorless and purple sulfur bacteria, during the oxidation of sulfides, mineral sulfur is deposited inside the cell in the form of drops. Sulfur accumulation occurs in environments rich in hydrogen sulfide H2S. When sulfides are depleted from the environment, bacteria use intracellular sulfur. For colorless sulfur bacteria it serves as a source of energy, for photosynthetic purple sulfur bacteria it serves as an electron donor.
In cyanobacteria, the reserve substance is cyanophycin. It is a polypeptide consisting of arginine and aspartic acid. It serves as a source of nitrogen when there is a lack of it in the environment. The accumulation of cyanophycin granules occurs in the stationary phase of culture growth and can amount to up to 8% of the dry weight of the cell.
The outer cytoplasmic membrane is a thin film. Its thickness is about 7-10 nm. The film is viewed only through an electron microscope.
Structure
What is the composition of the cytoplasmic membrane? The structure of the film is quite diverse. According to its chemical organization, it is a complex of proteins and lipids. The cytoplasmic membrane of a cell includes a bilayer. It acts as a base. In addition, the cytoplasmic membrane contains cholesterol and glycolipids. These substances are characterized by amphipatricity. In other words, they contain hydrophobic (“water-afraid”) and hydrophilic (“water-loving”) ends. The latter (phosphate group) are directed outward from the membrane, the latter (residues from fatty acids) are oriented towards each other. Due to this, a lipid bipolar layer is formed. Lipid molecules have mobility. They are able to move in their own monolayer or (rarely) from one to another.
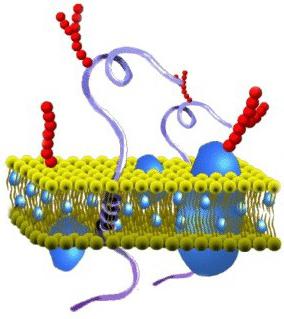
The lipid layer can be in a solid or liquid crystal state. Monolayers are asymmetrical. This means that their lipid composition is different. Due to this property, cytoplasmic membranes have specificity even within a single cell. The second essential component of the film includes proteins. Many of these connections can move in the membrane plane or rotate around their own axis. However, they are not able to move from one part of the bilayer to another. Protecting the internal environment is the main task performed by the cytoplasmic membrane. The structure of the film, in addition, ensures the course of various processes. Proteins are responsible for performing certain tasks. Lipids provide the structural features of the film.
Cytoplasmic membrane: functions
The main objectives are:
- Barrier. The protective film provides active, passive, selective, regulated exchange of compounds with the external environment. Due to selective permeability, the cell and its compartments are separated and supplied with the necessary substances.
- Transport. The transition of compounds from cell to cell occurs through the film. Thanks to this, nutritional compounds are delivered, metabolic end products are removed, and various substances are secreted. In addition, ionic gradients are formed, and ionic concentration and pH are maintained at optimal levels. They are necessary for the active activity of cell enzymes.
Auxiliary tasks

Special properties
The specific functions of the membrane include:

More information
If some particles, for one reason or another, are not able to pass through the phospholipid bilayer (for example, due to hydrophilic properties, since the cytoplasmic membrane inside is hydrophobic and does not allow such compounds to pass through, or due to the large size of the particles themselves), but they are necessary, then They can pass through with the help of special carrier proteins (transporters) and channel proteins. Or their penetration is carried out through endocytosis.

In the process of passive transport, substances cross the lipid layer by diffusion. In this case, no energy is wasted. Facilitated diffusion may be one of the variants of such a mechanism. During this process, a specific molecule facilitates the passage of the substance. It may have a channel capable of passing only particles of the same type. Active transport consumes energy. This is due to the fact that this process is carried out against a concentration gradient. The cytoplasmic membrane contains special pump proteins, including ATPase, which promotes the active entry of potassium ions and the excretion of sodium ions.
Models
There are several of them:
- "Sandwich model". The idea of a three-layer structure of all membranes was expressed by scientists Dawson and Danieli in 1935. In their opinion, the structure of the film was as follows: proteins-lipids-proteins. This idea existed for quite a long time.
- "Liquid-mosaic structure." This model was described by Nicholson and Singer in 1972. In accordance with it, protein molecules do not form a continuous layer, but are immersed in a bipolar lipid layer in the form of a mosaic to varying depths. This model is considered the most universal.
- "Protein crystal structure". According to this model, membranes are formed due to the interweaving of protein and lipid molecules, which are combined on the basis of hydrophilic-hydrophobic bonds.
Cell membrane also called plasma (or cytoplasmic) membrane and plasmalemma. This structure not only separates the internal contents of the cell from the external environment, but is also part of most cellular organelles and the nucleus, in turn separating them from the hyaloplasm (cytosol) - the viscous-liquid part of the cytoplasm. Let's agree to call cytoplasmic membrane the one that separates the contents of the cell from the external environment. The remaining terms denote all membranes.
The structure of the cellular (biological) membrane is based on a double layer of lipids (fats). The formation of such a layer is associated with the characteristics of their molecules. Lipids do not dissolve in water, but condense in it in their own way. One part of a single lipid molecule is a polar head (it is attracted to water, i.e. hydrophilic), and the other is a pair of long non-polar tails (this part of the molecule is repelled by water, i.e. hydrophobic). This structure of molecules causes them to “hide” their tails from the water and turn their polar heads towards the water.
The result is a lipid bilayer in which the nonpolar tails are inward (facing each other) and the polar heads are outward (toward the external environment and cytoplasm). The surface of such a membrane is hydrophilic, but inside it is hydrophobic.
In cell membranes, phospholipids predominate among the lipids (they belong to complex lipids). Their heads contain a phosphoric acid residue. In addition to phospholipids, there are glycolipids (lipids + carbohydrates) and cholesterol (related to sterols). The latter imparts rigidity to the membrane, being located in its thickness between the tails of the remaining lipids (cholesterol is completely hydrophobic).
Due to electrostatic interaction, some protein molecules are attached to the charged lipid heads, which become surface membrane proteins. Other proteins interact with nonpolar tails, are partially buried in the bilayer, or penetrate through it.
Thus, the cell membrane consists of a bilayer of lipids, surface (peripheral), embedded (semi-integral) and permeating (integral) proteins. In addition, some proteins and lipids on the outside of the membrane are associated with carbohydrate chains.

This fluid mosaic model of membrane structure was put forward in the 70s of the XX century. Previously, a sandwich model of structure was assumed, according to which the lipid bilayer is located inside, and on the inside and outside the membrane is covered with continuous layers of surface proteins. However, the accumulation of experimental data refuted this hypothesis.
The thickness of membranes in different cells is about 8 nm. Membranes (even different sides of one) differ from each other in the percentage of different types of lipids, proteins, enzymatic activity, etc. Some membranes are more liquid and more permeable, others are more dense.
Cell membrane breaks easily merge due to the physicochemical properties of the lipid bilayer. In the plane of the membrane, lipids and proteins (unless they are anchored by the cytoskeleton) move.
Functions of the cell membrane
Most proteins immersed in the cell membrane perform an enzymatic function (they are enzymes). Often (especially in the membranes of cell organelles) enzymes are located in a certain sequence so that the reaction products catalyzed by one enzyme move to the second, then the third, etc. A conveyor is formed that stabilizes surface proteins, because they do not allow the enzymes to float along the lipid bilayer.
The cell membrane performs a delimiting (barrier) function from the environment and at the same time transport functions. We can say that this is its most important purpose. The cytoplasmic membrane, having strength and selective permeability, maintains the constancy of the internal composition of the cell (its homeostasis and integrity).
In this case, the transport of substances occurs in various ways. Transport along a concentration gradient involves the movement of substances from an area with a higher concentration to an area with a lower concentration (diffusion). For example, gases (CO 2 , O 2 ) diffuse.
There is also transport against a concentration gradient, but with energy consumption.
Transport can be passive and facilitated (when it is helped by some kind of carrier). Passive diffusion across the cell membrane is possible for fat-soluble substances.
There are special proteins that make membranes permeable to sugars and other water-soluble substances. Such carriers bind to transported molecules and pull them through the membrane. This is how glucose is transported inside red blood cells.
Threading proteins combine to form a pore for the movement of certain substances across the membrane. Such carriers do not move, but form a channel in the membrane and work similarly to enzymes, binding a specific substance. Transfer occurs due to a change in protein conformation, resulting in the formation of channels in the membrane. An example is the sodium-potassium pump.
The transport function of the eukaryotic cell membrane is also realized through endocytosis (and exocytosis). Thanks to these mechanisms, large molecules of biopolymers, even whole cells, enter the cell (and out of it). Endo- and exocytosis are not characteristic of all eukaryotic cells (prokaryotes do not have it at all). Thus, endocytosis is observed in protozoa and lower invertebrates; in mammals, leukocytes and macrophages absorb harmful substances and bacteria, i.e. endocytosis performs a protective function for the body.
Endocytosis is divided into phagocytosis(cytoplasm envelops large particles) and pinocytosis(capturing droplets of liquid with substances dissolved in it). The mechanism of these processes is approximately the same. Absorbed substances on the surface of cells are surrounded by a membrane. A vesicle (phagocytic or pinocytic) is formed, which then moves into the cell.
Exocytosis is the removal of substances from the cell by the cytoplasmic membrane (hormones, polysaccharides, proteins, fats, etc.). These substances are contained in membrane vesicles that approach the cell membrane. Both membranes merge and the contents appear outside the cell.
The cytoplasmic membrane performs a receptor function. To do this, structures are located on its outer side that can recognize a chemical or physical stimulus. Some of the proteins that penetrate the plasmalemma are connected from the outside to polysaccharide chains (forming glycoproteins). These are peculiar molecular receptors that capture hormones. When a particular hormone binds to its receptor, it changes its structure. This in turn triggers the cellular response mechanism. In this case, channels can open, and certain substances can begin to enter or exit the cell.
The receptor function of cell membranes has been well studied based on the action of the hormone insulin. When insulin binds to its glycoprotein receptor, the catalytic intracellular part of this protein (adenylate cyclase enzyme) is activated. The enzyme synthesizes cyclic AMP from ATP. Already it activates or suppresses various enzymes of cellular metabolism.
The receptor function of the cytoplasmic membrane also includes recognition of neighboring cells of the same type. Such cells are attached to each other by various intercellular contacts.
In tissues, with the help of intercellular contacts, cells can exchange information with each other using specially synthesized low-molecular substances. One example of such an interaction is contact inhibition, when cells stop growing after receiving information that free space is occupied.
Intercellular contacts can be simple (the membranes of different cells are adjacent to each other), locking (invaginations of the membrane of one cell into another), desmosomes (when the membranes are connected by bundles of transverse fibers that penetrate the cytoplasm). In addition, there is a variant of intercellular contacts due to mediators (intermediaries) - synapses. In them, the signal is transmitted not only chemically, but also electrically. Synapses transmit signals between nerve cells, as well as from nerve to muscle cells.
The cytoplasmic cell membrane consists of three layers:
External – protein;
Middle - bimolecular layer of lipids;
Internal - protein.
Membrane thickness is 7.5-10 nm. The bimolecular layer of lipids is the matrix of the membrane. The lipid molecules of both layers interact with the protein molecules immersed in them. From 60 to 75% of membrane lipids are phospholipids, 15-30% are cholesterol. Proteins are represented mainly by glycoproteins. Distinguish integral proteins, permeating the entire membrane, and peripheral located on the outer or inner surface.
Integral proteins form ion channels that ensure the exchange of certain ions between extra- and intracellular fluid. They are also enzymes that carry out counter-gradient transport of ions across the membrane.
Peripheral proteins are chemoreceptors on the outer surface of the membrane that can interact with various physiologically active substances.
Membrane functions:
1. Ensures the integrity of the cell as a structural unit of tissue.
Carries out the exchange of ions between the cytoplasm and extracellular fluid.
Provides active transport of ions and other substances into and out of the cell.
Performs the perception and processing of information coming to the cell in the form of chemical and electrical signals.
Mechanisms of cell excitability. History of the study of bioelectric phenomena.
Most information transmitted in the body takes the form of electrical signals (for example, nerve impulses). The presence of animal electricity was first established by the natural scientist (physiologist) L. Galvani in 1786. In order to study atmospheric electricity, he suspended neuromuscular preparations of frog legs on a copper hook. When these paws touched the iron railings of the balcony, muscle contraction occurred. This indicated the action of some kind of electricity on the nerve of the neuromuscular drug. Galvani believed that this was due to the presence of electricity in the living tissues themselves. However, A. Volta established that the source of electricity is the place of contact of two dissimilar metals - copper and iron. In physiology Galvani's first classical experiment is considered to be touching the nerve of the neuromuscular preparation with bimetallic tweezers made of copper and iron. To prove he was right, Galvani produced second experience. He threw the end of the nerve innervating the neuromuscular preparation onto the cut of its muscle. As a result, it was reduced. However, this experience did not convince Galvani’s contemporaries. Therefore, another Italian, Matteuci, performed the following experiment. He superimposed the nerve of one frog neuromuscular preparation onto the muscle of the second, which contracted under the influence of an irritating current. As a result, the first drug also began to shrink. This indicated the transfer of electricity (action potential) from one muscle to another. The presence of a potential difference between the damaged and undamaged areas of the muscle was first accurately established in the 19th century using a string galvanometer (ammeter) by Matteuci. Moreover, the cut had a negative charge, and the surface of the muscle had a positive charge.