
Jaké vlastnosti má cytoplazmatická membrána? Buněčná membrána
Vnější cytoplazmatická membrána je tenký film. Jeho tloušťka je asi 7-10 nm. Film je pozorován pouze prostřednictvím elektronového mikroskopu.
Struktura
Jaké je složení cytoplazmatické membrány? Struktura filmu je poměrně různorodá. Podle chemického uspořádání se jedná o komplex proteinů a lipidů. Cytoplazmatická membrána buňky zahrnuje dvojvrstvu. Působí jako základna. Kromě toho cytoplazmatická membrána obsahuje cholesterol a glykolipidy. Tyto látky se vyznačují amfipatričností. Jinými slovy, obsahují hydrofobní („vody se bojí“) a hydrofilní („vodu milující“) konce. Posledně jmenované (fosfátová skupina) směřují ven z membrány, druhé (zbytky mastných kyselin) jsou orientovány proti sobě. Díky tomu se vytváří lipidová bipolární vrstva. Molekuly lipidů mají pohyblivost. Jsou schopni se pohybovat ve své vlastní monovrstvě nebo (zřídka) z jedné do druhé.

Lipidová vrstva může být v pevném nebo tekutém krystalovém stavu. Monovrstvy jsou asymetrické. To znamená, že jejich lipidové složení je odlišné. Díky této vlastnosti mají cytoplazmatické membrány specificitu i v rámci jedné buňky. Druhou podstatnou složkou filmu jsou bílkoviny. Mnoho z těchto spojení se může pohybovat v rovině membrány nebo se otáčet kolem své vlastní osy. Nejsou však schopni přecházet z jedné části dvojvrstvy do druhé. Ochrana vnitřního prostředí je hlavním úkolem cytoplazmatické membrány. Struktura filmu navíc zajišťuje průběh různých procesů. Proteiny jsou zodpovědné za provádění určitých úkolů. Lipidy poskytují strukturální vlastnosti filmu.
Cytoplazmatická membrána: funkce
Hlavní úkoly jsou:
- Bariéra. Ochranný film zajišťuje aktivní, pasivní, selektivní, regulovanou výměnu sloučenin s vnějším prostředím. Díky selektivní propustnosti jsou buňka a její kompartmenty odděleny a zásobovány potřebnými látkami.
- Doprava. K přechodu sloučenin z buňky do buňky dochází prostřednictvím filmu. Díky tomu jsou dodávány nutriční sloučeniny, odstraňovány konečné produkty metabolismu a vylučovány různé látky. Kromě toho se vytvářejí iontové gradienty a koncentrace iontů a pH jsou udržovány na optimálních úrovních. Jsou nezbytné pro aktivní činnost buněčných enzymů.
Pomocné úkoly

Speciální vlastnosti
Mezi specifické funkce membrány patří:

Více informací
Pokud některé částice z toho či onoho důvodu nejsou schopny projít fosfolipidovou dvojvrstvou (například kvůli hydrofilním vlastnostem, protože cytoplazmatická membrána uvnitř je hydrofobní a neumožňuje takovým sloučeninám projít, nebo kvůli velké velikost samotných částic), ale jsou nezbytné, pak mohou procházet pomocí speciálních nosných proteinů (transportérů) a kanálových proteinů. Nebo se jejich penetrace provádí endocytózou.
V procesu pasivního transportu látky procházejí lipidovou vrstvou difúzí. V tomto případě nedochází k plýtvání energií. Jednou z variant takového mechanismu může být usnadněná difúze. Během tohoto procesu specifická molekula usnadňuje průchod látky. Může mít kanál schopný procházet pouze částicemi stejného typu. Aktivní doprava spotřebovává energii. To je způsobeno skutečností, že tento proces se provádí proti koncentračnímu gradientu. Cytoplazmatická membrána obsahuje speciální pumpové proteiny včetně ATPázy, která podporuje aktivní vstup iontů draslíku a vylučování iontů sodíku.
Modelky
Je jich několik:
- "Sendvičový model". Myšlenku třívrstvé struktury všech membrán vyslovili vědci Dawson a Danieli v roce 1935. Podle jejich názoru byla struktura filmu následující: proteiny-lipidy-proteiny. Tato myšlenka existovala poměrně dlouho.
- "Struktura tekuté mozaiky." Tento model byl popsán Nicholsonem a Singerem v roce 1972. V souladu s ní molekuly proteinů netvoří souvislou vrstvu, ale jsou do různé hloubky ponořeny do bipolární lipidové vrstvy ve formě mozaiky. Tento model je považován za nejuniverzálnější.
- "Krystalová struktura proteinu". Membrány podle tohoto modelu vznikají díky prolínání proteinových a lipidových molekul, které se spojují na základě hydrofilně-hydrofobních vazeb.
Každá živá buňka je oddělena od prostředí tenkou membránou zvláštní struktury - cytoplazmatickou membránou (CPM). Eukaryota mají četné intracelulární membrány, které oddělují prostor organel od cytoplazmy, zatímco pro většinu prokaryot je CPM jedinou buněčnou membránou. U některých bakterií a archeí může pronikat do cytoplazmy a vytvářet výrůstky a záhyby různých tvarů.
CPM všech buněk jsou sestaveny podle jediného plánu a sestávají z fosfolipidů (obr. 3.5, A). U bakterií obsahují dvě mastné kyseliny, obvykle s 16-18 atomy uhlíku v řetězci a s nasycenou nebo jednou nenasycenou vazbou, spojené esterovou vazbou se dvěma hydroxylovými skupinami glycerolu. Složení mastných kyselin bakterií se může měnit v reakci na změny prostředí, zejména teplotu. Při poklesu teploty se zvyšuje množství nenasycených mastných kyselin ve složení fosfolipidů, což výrazně ovlivňuje tekutost membrány. Některé mastné kyseliny mohou být rozvětvené nebo mohou obsahovat cyklopropanový kruh. Třetí OH skupina glycerolu je napojena na zbytek kyseliny fosforečné a přes něj na hlavní skupinu. Hlavní skupiny fosfolipidů mohou mít u různých prokaryot různou chemickou povahu (fosfatidylethanolamin, fosfatidylglycerol, kardiolipin, fosfatidylserin, lecitin atd.), ale mají jednodušší strukturu než u eukaryot. Například při E. coli, jsou zastoupeny 75 % fosfatidylethanolaminem, 20 % fosfatidylglycerolem, zbytek tvoří kardiolipin (difosfatidylglycerol), fosfatidylserin a stopová množství dalších sloučenin. Jiné bakterie mají složitější typy membránových lipidů. Některé buňky tvoří glykolipidy, jako je monogalaktosyldiglycerid. Archaální membránové lipidy se liší od eukaryotických a bakteriálních. Místo mastných kyselin obsahují vyšší isoprenoidní alkoholy připojené ke glycerolu spíše jednoduchou než esterovou vazbou.
Rýže. 3.5.
A- fosfolipid; b- dvouvrstvá membrána
O O o O o o
Takové molekuly tvoří membránovou dvojvrstvu, kde hydrofobní části směřují dovnitř a hydrofilní části směrem ven, do prostředí a do cytoplazmy (obr. 3.5, Obr. b).Četné proteiny jsou zabudovány do dvojvrstvy nebo ji protínají a mohou difundovat v membráně a někdy vytvářet složité komplexy. Membránové proteiny mají řadu důležitých funkcí, mezi které patří přeměna a ukládání metabolické energie, regulace vstřebávání a uvolňování všech živin a metabolických produktů. Kromě toho rozpoznávají a přenášejí mnoho signálů odrážejících změny v prostředí a spouštějí odpovídající kaskádu reakcí vedoucích k buněčné odpovědi. Tuto organizaci membrán dobře vysvětluje model tekutých krystalů s mozaikou proloženou membránovými proteiny (obr. 3.6).

Rýže. 3.6.
Většina biologických membrán má tloušťku 4 až 7 nm. Buněčné membrány jsou jasně viditelné v transmisním elektronovém mikroskopu v kontrastu s těžkými kovy. Na elektronových mikrosnímcích vypadají jako třívrstvé útvary: dvě vnější tmavé vrstvy ukazují polohu polárních skupin lipidů a světlá střední vrstva ukazuje hydrofobní vnitřní prostor (obr. 3.7).
Další technikou pro studium membrán je získání čipů buněk zmrazených na teplotu kapalného dusíku a kontrast výsledných povrchů naprašováním těžkými kovy.
(platina, zlato, stříbro). Výsledné přípravky se prohlížejí pod rastrovacím elektronovým mikroskopem. V tomto případě je vidět povrch membrány a v ní obsažené mozaikové membránové proteiny, které neprocházejí membránou, ale jsou spojeny speciálními hydrofobními kotevními oblastmi s hydrofobní oblastí dvojvrstvy.

Rýže. 3.7.
CPM má vlastnost selektivní permeability, brání volnému pohybu většiny látek dovnitř a ven z buňky a také hraje významnou roli v buněčném růstu a dělení, pohybu a exportu povrchových a extracelulárních proteinů a sacharidů (exopolysacharidů). . Pokud je buňka umístěna do prostředí s vyšším nebo nižším osmotickým tlakem než uvnitř cytoplazmy, pak voda buňku opustí nebo do ní vstoupí. To odráží vlastnost vody vyrovnávat gradienty roztoku. V tomto případě se cytoplazma stahuje nebo expanduje (fenomén plazmolýzy/deplazmolýzy). Většina bakterií však v takových experimentech nemění svůj tvar kvůli přítomnosti tuhé buněčné stěny.
CPM reguluje tok živin a metabolitů. Přítomnost hydrofobní vrstvy tvořené membránovými lipidy zabraňuje průchodu jakýchkoli polárních molekul a makromolekul přes ni. Tato vlastnost umožňuje buňkám, které většinou existují ve zředěných roztocích, zachovat si užitečné makromolekuly a metabolické prekurzory. Buněčná membrána je také navržena tak, aby vykonávala transportní funkci. Prokaryota mají obvykle velké množství velmi specifických transportních systémů. Transport je nedílnou součástí celkové bioenergetiky buňky, která prostřednictvím CPM vytváří a využívá různé iontové gradienty k transportu látek a vytváření dalších gradientů nezbytných pro buňku. CPM hraje významnou roli v buněčném pohybu, růstu a dělení. Mnoho metabolických procesů je soustředěno v membráně prokaryot. Membránové proteiny plní důležité funkce: podílejí se na přeměně a ukládání energie, regulují vstřebávání a uvolňování všech živin a metabolických produktů, rozpoznávají a přenášejí signály o změnách prostředí.
Vnější cytoplazmatická membrána obklopující cytoplazmu každé buňky určuje její velikost a zajišťuje udržení významných rozdílů mezi buněčným obsahem a prostředím. Membrána slouží jako vysoce selektivní filtr, který udržuje rozdíl v koncentracích iontů na obou stranách membrány a umožňuje pronikání živin do buňky a odvádění odpadních produktů.
Všechny biologické membrány jsou sestavy lipidových a proteinových molekul držených pohromadě nekovalentními interakcemi. Molekuly lipidů a proteinů tvoří souvislou dvojitou vrstvu.
Lipidová dvojvrstva je hlavní strukturou membrány, která vytváří relativně nepropustnou bariéru pro většinu ve vodě rozpustných molekul.
Molekuly bílkovin jsou jakoby „rozpuštěny“ v lipidové dvojvrstvě. Prostřednictvím proteinů jsou vykonávány různé funkce membrány: některé z nich zajišťují transport určitých molekul do buňky nebo z buňky, jiné jsou enzymy a katalyzují reakce spojené s membránou a další zajišťují strukturální spojení mezi cytoskeletem a extracelulární matricí. nebo slouží jako receptory pro příjem a přeměnu chemických signálů z prostředí.
Důležitou vlastností biologických membrán je tekutost. Všechny buněčné membrány jsou pohyblivé tekuté struktury: většina jejich základních lipidových a proteinových molekul je schopna se poměrně rychle pohybovat v rovině membrány. Další vlastností membrán je jejich asymetrie: obě jejich vrstvy se liší složením lipidů a proteinů, což odráží funkční rozdíly jejich povrchů.
Funkce vnější cytoplazmatické membrány:
· bariéra - zajišťuje regulovaný, selektivní, pasivní a aktivní metabolismus s okolím. Selektivní permeabilita zajišťuje oddělení buňky a buněčných kompartmentů od okolí a dodává jim potřebné látky.
Transport - látky jsou transportovány do a z buňky přes membránu. Transport přes membrány zajišťuje: dodávání živin, odstraňování konečných produktů metabolismu, sekreci různých látek, vytváření iontových gradientů, udržování vhodného pH a koncentrace iontů v buňce, které jsou nezbytné pro fungování buněčných enzymů.
Částice, které z nějakého důvodu nejsou schopny procházet fosfolipidovou dvojvrstvou (například kvůli hydrofilním vlastnostem, protože membrána uvnitř je hydrofobní a nepropouští hydrofilní látky, nebo kvůli jejich velké velikosti), ale jsou nezbytné pro buňky, mohou pronikat membránou prostřednictvím speciálních nosných proteinů (transportérů) a kanálových proteinů nebo endocytózou.
Při pasivním transportu látky procházejí lipidovou dvojvrstvou bez spotřeby energie, difúzí. Variantou tohoto mechanismu je usnadněná difúze, při které specifická molekula pomáhá látce projít membránou. Tato molekula může mít kanál, který umožňuje průchod pouze jednomu typu látky.
Aktivní transport vyžaduje energii, protože probíhá proti koncentračnímu gradientu. Na membráně jsou speciální pumpové proteiny včetně ATPázy, které aktivně pumpují draselné ionty (K+) do buňky a pumpují z ní ionty sodíku (Na+).
· matrice - zajišťuje určitou vzájemnou polohu a orientaci membránových proteinů, jejich optimální interakci;
· mechanická - zajišťuje autonomii buňky, jejích intracelulárních struktur, jakož i spojení s jinými buňkami (ve tkáních). Buněčné stěny hrají hlavní roli při zajišťování mechanické funkce a u zvířat mezibuněčná látka.
· energie - při fotosyntéze v chloroplastech a buněčném dýchání v mitochondriích fungují v jejich membránách systémy přenosu energie, na kterých se podílejí i bílkoviny;
· receptor - některé proteiny sedící v membráně jsou receptory (molekuly, s jejichž pomocí buňka vnímá určité signály).
Například hormony cirkulující v krvi působí pouze na cílové buňky, které mají receptory odpovídající těmto hormonům. Neurotransmitery (chemické látky, které zajišťují vedení nervových vzruchů) se také vážou na speciální receptorové proteiny v cílových buňkách.
· enzymatické - membránové proteiny jsou často enzymy. Například plazmatické membrány buněk střevního epitelu obsahují trávicí enzymy.
· realizace tvorby a vedení biopotenciálů.
Pomocí membrány je v buňce udržována konstantní koncentrace iontů: koncentrace iontů K+ uvnitř buňky je mnohem vyšší než venku a koncentrace Na+ je mnohem nižší, což je velmi důležité, protože to zajišťuje udržování rozdílu potenciálů na membráně a generování nervového impulsu.
· značení buněk – na membráně jsou antigeny, které fungují jako markery – „štítky“, které umožňují buňku identifikovat. Jedná se o glykoproteiny (tj. proteiny s navázanými rozvětvenými oligosacharidovými postranními řetězci), které hrají roli „antén“. Kvůli nesčetným konfiguracím postranních řetězců je možné vytvořit specifický marker pro každý typ buňky. Pomocí markerů mohou buňky rozpoznat jiné buňky a jednat ve shodě s nimi, například při tvorbě orgánů a tkání. To také umožňuje imunitnímu systému rozpoznat cizí antigeny.
Každé lidské nebo zvířecí tělo se skládá z miliard buněk. Buňka je složitý mechanismus, který plní specifické funkce. Všechny orgány a tkáně se skládají z podjednotek.
Systém má cytoplazmatickou membránu, cytoplazmu, jádro a řadu organel. Jádro je odděleno od organel vnitřní membránou. Všechny dohromady poskytují život tkáním a také umožňují metabolismus.
Ve fungování hraje důležitou roli cytoplazmatické lemma neboli membrána.
Samotný název, vnější cytoplazmatická membrána, pochází z latinského membrana, nebo jinak kůže. Jedná se o oddělovač prostoru mezi buněčnými organismy.
Hypotéza konstrukce byla předložena již v roce 1935. V. Robertson v roce 1959 dospěl k závěru, že membránové pláště jsou uspořádány podle stejného principu.
Díky velkému množství nashromážděných informací získala dutina kapalný mozaikový model struktury. Nyní je považován za všeobecně uznávaný. Je to vnější cytoplazmatická membrána, která tvoří vnější obal jednotek.
Struktura
Co je tedy plazmatické lemma?
Je to tenký film oddělující prokaryota od vnitřního prostředí. Lze to vidět pouze mikroskopem. Struktura cytoplazmatické membrány zahrnuje dvojvrstvu, která slouží jako základ.
Dvojvrstva je dvojitá vrstva skládající se z proteinů a lipidů. Existují také cholesterol a glykolipidy, které jsou amfipatické.
co to znamená?
Tučný organismus má bipolární hlavu a hydrofilní ocas. První je kvůli strachu z vody a druhý kvůli její absorpci. Skupina fosfátů má z filmu směr ven, ty jsou nasměrovány k sobě.
Tak se vytvoří bipolární lipidová vrstva. Lipidy jsou vysoce aktivní, mohou se pohybovat ve své monovrstvě a zřídka se přesouvají do jiných oblastí.
Polymery se dělí na:
- externí,
- integrální,
- prostupující plazmatické lemma.
První jsou umístěny pouze na povrchové části sinusu. Jsou drženy pohromadě elektrostatikou s bipolárními hlavami lipidových prvků. Zachovává nutriční enzymy. Integrální uvnitř, jsou zabudovány do samotné struktury pláště, spoje mění své umístění v důsledku pohybu eukaryot. Slouží jako jakýsi dopravník, postavený tak, že po nich proudí substráty a reakční produkty. Proteinové sloučeniny prostupující makrodutinou mají vlastnosti vytváření pórů pro vstup živin do těla.
Jádro
Jakákoli jednotka má jádro, to je její základ. Cytoplazmatická membrána má také organelu, jejíž struktura bude popsána níže.
Jaderná struktura zahrnuje membránu, mízu, místo sestavení ribozomu a chromatin. Plášť je rozdělen jaderným prostorem, je obklopený kapalinou.
Funkce organely jsou rozděleny do dvou hlavních:
- uzavření struktury v organele,
- regulace obsahu jádra a kapaliny.
Jádro se skládá z pórů, z nichž každý je určen přítomností kombinací těžkých pórů. Jejich objem může ukazovat na aktivní motorickou schopnost eukaryot. Například vysoce aktivní nezralé produkty obsahují větší počet oblastí pórů. Bílkoviny slouží jako jaderná šťáva.
Polymery představují kombinaci matrice a nukleoplazmy. Kapalina je obsažena uvnitř jaderného filmu a zajišťuje funkčnost genetického obsahu organismů. Proteinový prvek poskytuje ochranu a sílu podjednotkám.
Ribozomální RNA dozrávají v samotném jadérku. Samotné RNA geny jsou umístěny ve specifické oblasti několika chromozomů. V jejich rámci se tvoří malí organizátoři. Uvnitř se vytvářejí samotná jadérka. Zóny v mitotických chromozomech jsou reprezentovány konstrikcemi, nazývanými sekundární konstrikce. Při elektronickém vyšetření se rozlišují fáze vláknitého a granulačního původu.
Vývoj jádra
Další označení je fibrilární, pochází z proteinu a obrovských polymerů – předchozích verzí r-RNA. Následně tvoří menší prvky zralé rRNA. Když fibrila dozrává, stává se granulární strukturou nebo ribonukleoproteinovými granulemi.
Chromatin obsažený ve struktuře má barvicí vlastnosti. Je přítomen v nukleoplazmě jádra a slouží jako forma mezifáze pro životně důležitou aktivitu chromozomů. Složením chromatinu jsou řetězce DNA a polymery. Společně tvoří komplex nukleoproteinů.
Histony plní funkce organizování prostoru ve struktuře molekuly DNA. Kromě toho chromozomy zahrnují organické látky, enzymy obsahující polysacharidy a kovové částice. Chromatin se dělí na:
- euchromatin,
- heterochromatin.
První je způsobena nízkou hustotou, takže z takových eukaryot nelze číst genetická data.
Druhá možnost má kompaktní vlastnosti.
Struktura
Složení samotné skořápky je heterogenní. Díky neustálým pohybům se na něm objevují výrůstky a vybouleniny. Uvnitř je to kvůli pohybům makromolekul a jejich výstupu do jiné vrstvy.
Samotné látky vstupují dvěma způsoby:
- fagocytóza,
- pinocytóza.
Fagocytóza se projevuje invaginací pevných částic. Pinocytóza se týká výdutí. Vysunutím se okraje regionů přiblíží k sobě a zachytí tekutinu mezi eukaryoty.
Pinocytóza poskytuje mechanismus pro pronikání sloučenin do membrány. Průměr vakuoly se pohybuje od 0,01 do 1,3 um. Dále vakuola začne klesat do cytoplazmatické vrstvy a šněrovat se. Spojení mezi bublinami hraje roli transportu užitečných částic a rozkladu enzymů.
Trávicí cyklus
Celý okruh trávicí funkce je rozdělen do následujících fází:
- vstup složek do těla,
- rozklad enzymů
- vstup do cytoplazmy
- vylučování.
První fáze zahrnuje vstup látek do lidského těla. Poté se začnou rozkládat pomocí lysozomů. Oddělené částice pronikají do cytoplazmatického pole. Nestrávené zbytky jednoduše vycházejí přirozeně. Následně se sinus stane hustým a začne se přeměňovat na zrnité granule.
Membránové funkce
Jaké funkce tedy plní?
Hlavní budou:
- ochranný,
- přenosný,
- mechanický,
- matice,
- přenos energie,
- receptor.
Ochrana je vyjádřena jako bariéra mezi podjednotkou a vnějším prostředím. Film slouží jako regulátor výměny mezi nimi. V důsledku toho může být ten druhý aktivní nebo pasivní. Dochází k selektivitě potřebných látek.
V transportní funkci jsou spojení přenášena z jednoho mechanismu do druhého přes plášť. Právě tento faktor ovlivňuje dodávání užitečných sloučenin, odstraňování metabolických a rozkladných produktů a sekrečních složek. Vznikají gradienty iontové povahy, díky nimž je udržováno pH a úroveň koncentrace iontů.
Poslední dvě mise jsou pomocné. Práce na úrovni matrice je zaměřena na správné umístění proteinového řetězce uvnitř dutiny a jejich správné fungování. Díky mechanické fázi je článek zajištěn v autonomním režimu.
K přenosu energie dochází v důsledku fotosyntézy v zelených plastidech a respiračních procesů v buňkách uvnitř dutiny. Do práce se zapojují i bílkoviny. Proteiny díky své přítomnosti v membráně poskytují makrobuňce schopnost vnímat signály. Impulzy se pohybují z jedné cílové buňky do ostatních.
Mezi speciální vlastnosti membrány patří generování a implementace biopotenciálu, rozpoznávání buněk, a tedy značení.
Cytoplazmatická membrána oddělující cytoplazmu od buněčné stěny se nazývá plasmalemma (plazmatická membrána) a oddělující ji od vakuoly se nazývá tonoplast (elementární membrána).
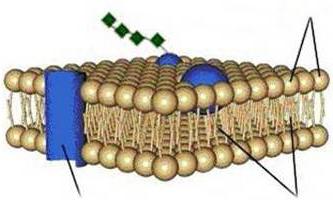
V současné době používají model membrány v tekuté mozaice (obr. 1.9), podle kterého se membrána skládá z dvojvrstvy lipidových molekul (fosfolipidů) s hydrofilními hlavami a 2 hydrofobními ocasy směřujícími dovnitř vrstvy. Kromě lipidů obsahují membrány také proteiny.
V bilipidové vrstvě se „vznášejí“ 3 typy membránových proteinů: integrální proteiny, které pronikají celou tloušťkou dvojvrstvy; polointegrální, neúplně pronikající dvojvrstvou; periferní, připojené z vnější nebo vnitřní strany membrány k jiným membránovým proteinům. Membránové proteiny plní různé funkce: některé z nich jsou enzymy, jiné působí jako nosiče specifických molekul přes membránu nebo tvoří hydrofilní póry, kterými mohou procházet polární molekuly.
Jednou z hlavních vlastností buněčných membrán je jejich polopropustnost: propouštějí vodu, ale nepropouštějí látky v ní rozpuštěné, tedy mají selektivní permeabilitu.
Rýže. 1.9. Schéma struktury biologické membrány:
A - extracelulární prostor; B - cytoplazma; 1 - bimolekulární vrstva lipidů; 2 - periferní protein; 3 - hydrofilní oblast integrálního proteinu; 4 - hydrofobní oblast integrálního proteinu; 5 - sacharidový řetězec
Transport přes membrány
V závislosti na energetickém výdeji se transport látek a iontů membránou dělí na pasivní, které nevyžadují energii, a aktivní, spojené se spotřebou energie. Pasivní transport zahrnuje procesy, jako je difúze, usnadněná difúze a osmóza.
Difúze je proces pronikání molekul přes lipidovou dvojvrstvu podél koncentračního gradientu (z oblasti s vyšší koncentrací do oblasti s nižší koncentrací). Čím menší je molekula a čím je nepolární, tím rychleji difunduje membránou.
S usnadněnou difúzí napomáhá určitý transportní protein průchodu látky přes membránu. Do buňky se tak dostávají různé polární molekuly, jako jsou cukry, aminokyseliny, nukleotidy atd.
Osmóza je difúze vody přes polopropustné membrány. Osmóza způsobuje pohyb vody z roztoku s vysokým vodním potenciálem do roztoku s nízkým vodním potenciálem.
Aktivní doprava- jedná se o přenos molekul a iontů přes membránu, doprovázený energetickými náklady. Aktivní transport jde proti koncentračnímu gradientu a elektrochemickému gradientu a využívá energii ATP. Mechanismus aktivního transportu látek je založen na práci protonové pumpy (H+ a K+) u rostlin a hub, které uvnitř buňky udržují vysokou koncentraci K+ a nízkou koncentraci H+ (u živočichů Na+ a K+). Energie potřebná k provozu této pumpy je dodávána ve formě ATP, syntetizovaného během buněčného dýchání.
Je znám další typ aktivního transportu – endo- a exocytóza. Jedná se o 2 aktivní procesy, kterými jsou různé molekuly transportovány přes membránu do buňky ( endocytóza) nebo z něj ( exocytóza).
Při endocytóze se látky dostávají do buňky v důsledku invaginace (invaginace) plazmatické membrány. Vzniklé vezikuly neboli vakuoly jsou transportovány do cytoplazmy spolu s látkami v nich obsaženými. Absorpce velkých částic, jako jsou mikroorganismy nebo zbytky buněk, se nazývá fagocytóza. V tomto případě se tvoří velké bubliny zvané vakuoly. Absorpce kapalin (suspenzí, koloidních roztoků) nebo rozpuštěných látek pomocí malých bublinek je tzv. pinocytóza.
Reverzní proces endocytózy se nazývá exocytóza. Mnoho látek je z buňky odstraněno ve speciálních vezikulách nebo vakuolách. Příkladem je stažení jejich tekutých sekretů ze sekrečních buněk; dalším příkladem je účast dictyosomových vezikul na tvorbě buněčné stěny.
PROTOPLASTOVÉ DERIVÁTY
Vacuole
Vacuole- Toto je rezervoár ohraničený jedinou membránou - tonoplastem. Vakuola obsahuje buněčnou mízu - koncentrovaný roztok různých látek, jako jsou minerální soli, cukry, pigmenty, organické kyseliny, enzymy. Ve zralých buňkách se vakuoly spojují v jednu, centrální.
Vakuoly uchovávají různé látky, včetně konečných produktů metabolismu. Osmotické vlastnosti buňky silně závisí na obsahu vakuoly.
Vzhledem k tomu, že vakuoly obsahují silné roztoky solí a dalších látek, rostlinné buňky neustále osmoticky absorbují vodu a vytvářejí hydrostatický tlak na buněčnou stěnu, tzv. turgorový tlak. Proti turgorovému tlaku působí stejný tlak z buněčné stěny směřující do buňky. Většina rostlinných buněk existuje v hypotonickém prostředí. Ale pokud je taková buňka umístěna do hypertonického roztoku, voda začne buňku opouštět podle zákonů osmózy (aby se vyrovnal vodní potenciál na obou stranách membrány). Vakuola se zmenší, její tlak na protoplast se sníží a membrána se začne vzdalovat od buněčné stěny. Jev oddělení protoplastů od buněčné stěny se nazývá plazmolýza. V přirozených podmínkách taková ztráta turgoru v buňkách povede k uschnutí rostliny, povadnutí listů a stonků. Tento proces je však vratný: pokud je buňka umístěna do vody (např. při zalévání rostliny), dochází k jevu, který je opakem plazmolýzy – deplazmolýze (viz obr. 1.10).
Rýže. 1.10. Schéma plazmolýzy:
A - buňka ve stavu turgoru (v izotonickém roztoku); B - začátek plazmolýzy (buňka umístěna v 6% roztoku KNO3); B - úplná plazmolýza (buňka umístěna v 10% roztoku KNO3); 1 - chloroplast; 2 - jádro; 3 - buněčná stěna; 4 - protoplast; 5 - centrální vakuola
Inkluze
Buněčné inkluze jsou zásobní a vylučovací látky.
Rezervní látky (dočasně vyloučené z metabolismu) a s nimi odpadní (vylučovací látky) se často nazývají ergastickými látkami buňky. Zásobní látky zahrnují zásobní bílkoviny, tuky a sacharidy. Tyto látky se během vegetace hromadí v semenech, plodech, podzemních orgánech rostlin a v jádru stonku.
Náhradní látky
Zásobní bílkoviny, příbuzné jednoduchým bílkovinám – bílkovinám, se často ukládají v semenech. Vysrážené proteiny ve vakuolách tvoří kulatá nebo eliptická zrna nazývaná aleuron. Pokud aleuronová zrna nemají rozeznatelnou vnitřní strukturu a jsou složena z amorfního proteinu, nazývají se jednoduchá. Pokud se v zrnech aleuronu nachází mezi amorfním proteinem krystalická struktura (krystaloid) a lesklá bezbarvá tělíska kulatého tvaru (globoidy), nazýváme taková zrna aleuronu komplexní (viz obr. 1.11). Amorfní protein zrna aleuronu je homogenní, neprůhledný, nažloutlý protein, který bobtná ve vodě. Krystaloidy mají charakteristický romboedrický tvar krystalů, ale na rozdíl od pravých krystalů jejich bílkovina ve vodě bobtná. Globoidy se skládají z vápenato-hořečnaté soli, obsahují fosfor, jsou nerozpustné ve vodě a nereagují s bílkovinami.
Rýže. 1.11. Komplexní aleuronová zrna:
1 - póry ve skořápce; 2 - globoidy; 3 - amorfní proteinová hmota; 4 - krystaloidy ponořené do amforové proteinové hmoty
Zásobní lipidy obvykle se nacházejí v hyaloplazmě ve formě kapiček a nacházejí se téměř ve všech rostlinných buňkách. Toto je hlavní typ rezervních živin ve většině rostlin: semena a plody jsou na ně nejbohatší. Tuky (lipidy) jsou nejvíce kalorickou rezervní látkou. Činidlo pro látky podobné tuku je Sudan III, které je barví do oranžova.
Sacharidy jsou obsaženy ve složení každé buňky ve formě ve vodě rozpustných cukrů (glukóza, fruktóza, sacharóza) a ve vodě nerozpustných polysacharidů (celulóza, škrob). Sacharidy hrají v buňce roli zdroje energie pro metabolické reakce. Cukry po vazbě s jinými biologickými látkami buňky tvoří glykosidy a polysacharidy s bílkovinami glykoproteiny. Složení sacharidů v rostlinné buňce je mnohem rozmanitější než v živočišných buňkách, a to díky různorodému složení polysacharidů buněčné stěny a cukrů v buněčné míze vakuol.
Hlavním a nejběžnějším zásobním sacharidem je polysacharid škrob. Primární asimilační škrob se tvoří v chloroplastech. V noci, kdy ustává fotosyntéza, je škrob hydrolyzován na cukry a transportován do zásobních tkání – hlíz, cibulí, oddenků. Tam se ve speciálních typech leukoplastů – amyloplastech – část cukrů ukládá ve formě zrnek sekundárního škrobu. Škrobová zrna se vyznačují vrstvením, které se vysvětluje rozdílným obsahem vody v důsledku nerovnoměrného přísunu škrobu během dne. V tmavých vrstvách je více vody než ve světlých. Zrno s jedním středem tvorby škrobu ve středu amyloplastu se nazývá jednoduché soustředné, pokud je střed posunuto, nazývá se jednoduché excentrické. Zrno s několika centry tvořícími škrob je složité. V semisložených zrnech se nové vrstvy ukládají kolem několika škrobotvorných center a poté se tvoří společné vrstvy a pokrývají škrobotvorná centra (viz obr. 1.12). Činidlo pro škrob je roztok jódu, který dává modrou barvu.
Rýže. 1.12. Zrna bramborového škrobu (A):
1 - jednoduché zrno; 2 - polokomplex; 3 - komplexní; pšenice (B), oves (C)
Vylučovací látky (sekundární metabolické produkty)
Mezi buněčné inkluze patří také vylučovací látky, například krystaly šťavelanu vápenatého ( monokrystaly, raphidy - jehličkovité krystaly, drúzy - krystalové srůsty, krystalický písek - nahromadění mnoha malých krystalů) (viz obr. 1.13). Méně často jsou krystaly složeny z uhličitanu vápenatého nebo oxidu křemičitého ( cystolity; viz obr. 1.14). Na buněčné stěně se ukládají cystolity, vyčnívající do buňky v podobě hroznů a jsou charakteristické např. pro zástupce čeledi kopřivovitých, listy fíkusu.
Na rozdíl od živočichů, kteří přebytečné soli vylučují močí, rostliny nemají vyvinuté vylučovací orgány. Proto se má za to, že krystaly šťavelanu vápenatého jsou konečným produktem metabolismu protoplastů, vytvořeným jako zařízení pro odstraňování přebytečného vápníku z metabolismu. Tyto krystaly se zpravidla hromadí v orgánech, které rostlina periodicky zbavuje (listy, kůra).

Rýže. 1.13. Formy krystalů šťavelanu vápenatého v buňkách:
1, 2 - raphida (impatiens; 1 - boční pohled, 2 - řez); 3 - drúza (opuncie); 4 - krystalický písek (brambory); 5 - monokrystal (vanilka)
Rýže. 1.14. Cystolith (na průřezu listu fíkusu):
1 - listová slupka; 2 - cystolitida
Éterické oleje se hromadí v listech (máta, levandule, šalvěj), květech (šípky), plodech (citrusy) a semenech rostlin (kopr, anýz). Éterické oleje se nepodílejí na metabolismu, ale jsou široce používány v parfumerii (růžové, jasmínové oleje), potravinářském průmyslu (anýz, kopr), lékařství (máta, eukalyptové oleje). Rezervoáry pro akumulaci éterických olejů mohou být žlázy (máta), lysogenní nádobky (citrusové plody), žlázové chloupky (pelargónie).
Pryskyřice- jedná se o komplexní sloučeniny vznikající během normálního života nebo v důsledku destrukce tkáně. Jsou tvořeny epiteliálními buňkami vystýlajícími pryskyřičné kanálky jako vedlejší produkt metabolismu, často s éterickými oleji. Mohou se hromadit v buněčné míze, cytoplazmě ve formě kapek nebo v nádobách. Jsou nerozpustné ve vodě, nepropustné pro mikroorganismy a díky svým antiseptickým vlastnostem zvyšují odolnost rostlin vůči chorobám. Pryskyřice se používají v lékařství, ale i při výrobě barev, laků a mazacích olejů. V moderním průmyslu je nahrazují syntetické materiály.
Buněčná stěna
Tuhá buněčná stěna obklopující buňku sestává z celulózových mikrofibril uložených v matrici obsahující hemicelulózy a pektinové látky. Buněčná stěna poskytuje mechanickou podporu buňce, chrání protoplast a udržuje tvar buňky. V tomto případě je buněčná stěna schopna protažení. Stěna, která je produktem životně důležité činnosti protoplastu, může růst pouze v kontaktu s ním. Buněčnou stěnou se pohybuje voda a minerální soli, ta je však pro vysokomolekulární látky zcela nebo částečně nepropustná. Když protoplast odumře, stěna může nadále plnit funkci vedení vody. Přítomnost buněčné stěny, více než všechny ostatní vlastnosti, odlišuje rostlinné buňky od živočišných. Architektura buněčné stěny je do značné míry určena celulózou. Monomerem celulózy je glukóza. Svazky molekul celulózy tvoří micely, které se spojují do větších svazků – mikrofibril. Činidlo pro celulózu je chlor-zinek-jod (Cl-Zn-I), který dává modrofialové zbarvení.
Celulózová struktura buněčné stěny je vyplněna molekulami necelulózové matrice. Matrice obsahuje polysacharidy zvané hemicelulózy; pektinové látky (pektin), velmi blízké hemicelulózám, a glykoproteiny. Pektické látky, splývající mezi sousedními buňkami, tvoří střední desku, která se nachází mezi primárními membránami sousedních buněk. Při rozpuštění nebo zničení střední desky (k čemuž dochází v dužině zralých plodů) dochází k maceraci (z latinského maceratio – měknutí). Přirozenou maceraci lze pozorovat u mnoha přezrálých plodů (meloun, meloun, broskev). Umělá macerace (kdy je tkáň ošetřena zásadou nebo kyselinou) se používá k přípravě různých anatomických a histologických preparátů.
Buněčná stěna v procesu života může podléhat různým modifikacím - lignifikaci, suberizaci, slizu, kutinizaci, mineralizaci (viz tab. l.4).
Tabulka 1.4.
Související informace.