
Каким свойством обладает цитоплазматическая мембрана. Клеточная мембрана
Наружная цитоплазматическая мембрана представляет собой тончайшую пленку. Ее толщина - порядка 7-10 нм. Просматривается пленка только в электронный микроскоп.
Структура
Какой состав имеет цитоплазматическая мембрана? Строение пленки достаточно разнообразно. В соответствии с химической организацией, она представляет собой комплекс белков и липидов. Цитоплазматическая мембрана клетки включает в себя бислой. Он выступает в качестве основы. Кроме этого, цитоплазматическая мембрана содержит холестерол и гликолипиды. Этим веществам свойственна амфипатричность. Другими словами, в них присутствуют гидрофобные ("боящиеся влаги") и гидрофильные ("любящие воду") концы. Последние (фосфатная группа) направлены наружу от мембраны, вторые (остатки от жирных кислот) ориентированы друг к другу. За счет этого и формируется липидный биполярный слой. Липидные молекулы обладают подвижностью. Они способны перемещаться в собственном монослое либо (что редко) из одного в другой.

Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды - основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Цитоплазматическая мембрана: функции
Основными задачами являются:
- Барьерная . Защитная пленка обеспечивает активный, пассивный, избирательный, регулируемый обмен соединений с внешней средой. За счет избирательной проницаемости осуществляется отделение клетки и ее компартментов и снабжение их нужными веществами.
- Транспортная . Сквозь пленку осуществляется переход соединений от клетки к клетке. Благодаря этому доставляются питательные соединения, удаляются конечные продукты обмена, происходит секреция разных веществ. Кроме этого, формируются ионные градиенты, на оптимальном уровне поддерживаются ионная концентрация и рН. Они необходимы для активной деятельности ферментов клетки.
Вспомогательные задачи

Особые свойства
К специфическим функциям мембраны относят:

Дополнительные сведения
Если какие-то частицы по тем или другим причинам не способны пройти сквозь фосфолипидный бислой (к примеру, вследствие гидрофильных свойств, поскольку внутри цитоплазматическая мембрана гидрофобна и такие соединения не пропускает, либо из-за больших размеров самих частиц), но они необходимы, то пройти они могут с помощью специальных белков-переносчиков (транспортеров) и белков-каналов. Либо проникновение их осуществляется посредством эндоцитоза.
В процессе пассивного транспорта пересечение веществами липидного слоя происходит путем диффузии. При этом энергия не затрачивается. В качестве одного из вариантов такого механизма может выступать облегченная диффузия. В ходе нее облегчает прохождение вещества какая-нибудь специфическая молекула. У нее может присутствовать канал, способный пропускать только однотипные частицы. При активном транспорте затрачивается энергия. Это связано с тем, что данный процесс осуществляется против концентрационного градиента. Цитоплазматическая мембрана содержит особые белки-насосы, АТФазу в том числе, которая способствует активному вхождению калиевых и выведению натриевых ионов.
Модели
Их существует несколько:
- "Бутербродная модель" . Идею о трехслойном строении всех мембран высказали ученые Даусон и Даниэли в 1935 году. По их мнению, структура пленки была следующей: белки-липиды-белки. Такое представление существовало достаточно долго.
- "Жидкостно-мозаичная структура". Эта модель была описана Николсоном и Сингером в 1972 году. В соответствии с ней белковые молекулы не формируют сплошной слой, а погружаются в биполярный липидный в виде мозаики на различную глубину. Эта модель считается наиболее универсальной.
- "Белково-кристаллическая структура". В соответствии с этой моделью мембраны формируются за счет переплетения белковых и липидных молекул, которые объединены на базе гидрофильно-гидрофобных связей.
Любая живая клетка отделена от окружающей среды тонкой оболочкой особого строения - цитоплазматической мембраной (ЦПМ). Эукариоты имеют многочисленные внутриклеточные мембраны, отделяющие пространство органелл от цитоплазмы, тогда как для большинства прокариот ЦПМ - это единственная мембрана клетки. У некоторых бактерий и архей она может внедряться внутрь цитоплазмы, образуя выросты и складки различной формы.
ЦПМ любых клеток построены по единому плану и состоят из фосфолипидов (рис. 3.5, а). У бактерий в их состав входят две жирные кислоты обычно с 16-18 атомами углерода в цепочке и с насыщенными или одной ненасыщенной связями, соединенные сложноэфирной связью с двумя гидроксильными группами глицерола. Состав жирных кислот бактерий может варьировать в ответ на изменения окружающей среды, в особенности, температуры. При понижении температуры в составе фосфолипидов увеличивается количество ненасыщенных жирных кислот, что в значительной степени отражается на текучести мембраны. Некоторые жирные кислоты могут быть разветвленными или содержать циклопропановое кольцо. Третья ОН-группа глицерола связана с остатком фосфорной кислоты и через него - с головной группой. Головные группы фосфолипидов могут у разных прокариот иметь разную химическую природу (фосфатидилэта- ноламин, фосфатидилглицерол, кардиолипин, фосфатидилсерин, лецитин и др.), но они устроены проще, чем у эукариот. Например, у Е. coli, они представлены на 75% фосфатидилэтаноламином, на 20% - фосфатидилглицеро- лом, остальные состоят из кардиолипина (дифосфатидилглицерола), фос- фатидилсерина и следовых количеств других соединений. Прочие бактерии имеют более сложные типы мембранных липидов. Некоторые клетки образуют гликолипиды, такие как моногалактозилдиглицерид. Мембранные липиды архей отличаются от эукариотических и бактериальных. Вместо жирных кислот у них присутствуют высшие изопреноидные спирты, прикрепленные к глицеролу простой, а не сложной эфирной связью.
Рис. 3.5.
а - фосфолипида; б - бислойной мембраны
О О о О о о
Такие молекулы составляют мембранный бислой, где гидрофобные части обращены вовнутрь, а гидрофильные - наружу, в окружающую среду и в цитоплазму (рис. 3.5, б). В бислой погружены или пересекают его многочисленные белки, которые могут диффундировать внутри мембраны, иногда образуя сложные комплексы. Мембранные белки имеют ряд важных функций, включая преобразование и запасание метаболической энергии, регуляцию поглощения и выброса всех питательных веществ и продуктов метаболизма. Кроме того, они узнают и передают многие сигналы, отражающие изменения в окружающей среде, и запускают соответствующий каскад реакций, приводящий к клеточному ответу. Такая организация мембран хорошо объясняется жидкокристаллической моделью с мозаичным вкраплением мембранных белков (рис. 3.6).

Рис. 3.6.
Большинство биологических мембран имеют толщину от 4 до 7 нм. Клеточные мембраны хорошо различимы в просвечивающем электронном микроскопе при контрастировании тяжелыми металлами. На электронных микрофотографиях они имеют вид трехслойных образований: два внешних темных слоя показывают положение полярных групп липидов, а светлый средний слой - гидрофобное внутреннее пространство (рис. 3.7).
Другая техника исследования мембран заключается в получении сколов замороженных при температуре жидкого азота клеток и контрастировании образующихся поверхностей с помощью напыления тяжелых металлов
(платина, золото, серебро). Полученные препараты просматривают в сканирующем электронном микроскопе. При этом можно увидеть поверхность мембраны и включенные в нее мозаично мембранные белки, которые не простираются сквозь мембрану, а связаны специальными гидрофобными якорными областями с гидрофобной областью бислоя.

Рис. 3.7.
ЦПМ обладает свойством избирательной проницаемости, препятствуя свободному продвижению большинства веществ внутрь и из клетки, а также играет значительную роль в росте и делении клеток, движении, экспорте поверхностных и внеклеточных белков и углеводов (экзополисахаридов). Если клетку поместить в среду с более высоким или более низким осмотическим давлением, чем внутри цитоплазмы, то произойдет выход воды из клетки или вход воды в нее. Это отражает свойство воды уравнивать градиенты растворов. Цитоплазма при этом сжимается или расширяется (явление плазмолиза/деплазмолиза). Большинство бактерий, однако, не меняет свою форму в таких экспериментах вследствие наличия ригидной клеточной стенки.
ЦПМ регулирует потоки питательных веществ и метаболитов. Наличие гидрофобного слоя, образованного мембранными липидами, препятствует прохождению через нее любых полярных молекул и макромолекул. Это свойство позволяет клеткам, существующим в большинстве случаев в разбавленных растворах, удерживать полезные макромолекулы и метаболические предшественники. Мембрана клетки призвана осуществлять и транспортную функцию. Обычно прокариоты имеют большое количество очень специфических транспортных систем. Транспорт - интегральная часть общей биоэнергетики клетки, которая создает и использует различные ионные градиенты через ЦПМ для переноса веществ и формирования других необходимых клетке градиентов. ЦПМ играет значительную роль в движении, росте и делении клеток. В мембране прокариот сосредоточены многие метаболические процессы. Мембранные белки выполняют важные функции: участвуют в преобразовании и запасании энергии, регулируют поглощение и выброс всех питательных веществ и продуктов метаболизма, узнают и передают сигналы об изменениях в окружающей среде.
Наружная цитоплазматическая мембрана, окружающая цитоплазму каждой клетки, определяет ее величину и обеспечивает сохранение существенных различий между клеточным содержимым и окружающей средой. Мембрана служит высокоизбирательным фильтром, который поддерживает разницу концентраций ионов по обе стороны мембраны и позволяет питательным веществам проникать внутрь клетки, а продуктам выделения выходить наружу.
Все биологические мембраны представляют собой ансамбли липидных и белковых молекул, удерживаемых вместе с помощью нековалентных взаимодействий. Липидные и белковые молекулы образуют непрерывный двойной слой.
Липидный бислой - это основная структура мембраны, которая создает относительно непроницаемый барьер для большинства водорастворимых молекул.
Белковые молекулы как бы «растворены» в липидном бислое. При посредстве белков выполняются разнообразные функции мембраны: одни из них обеспечивают транспорт определенных молекул внутрь клетки или из нее, другие являются ферментами и катализируют ассоциированные с мембраной реакции, а третьи осуществляют структурную связь цитоскелета с внеклеточным матриксом или служат рецепторами для получения и преобразования химических сигналов из окружающей среды.
Важное свойство биологических мембран - текучесть. Все клеточные мембраны представляют собой подвижные текучие структуры: большая часть составляющих их молекул липидов и белков способна достаточно быстро перемещаться в плоскости мембраны. Другое свойство мембран - их асимметрия: оба их слоя различаются по липидному и белковому составам, что отражает функциональные различия их поверхностей.
Функции наружной цитоплазматической мембраны:
· барьерная - обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
· транспортная - через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке соответствующего pH и ионной концентрации, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине не способные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза.
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии, путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивают в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+).
· матричная - обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие;
· механическая - обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных - межклеточное вещество.
· энергетическая - при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
· рецепторная - некоторые белки, сидящие в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
· ферментативная - мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
· осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
· маркировка клетки - на мембране есть антигены, действующие как маркеры - «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Каждый организм человека, либо животного состоит из миллиардов клеток. Клетка представляет собой сложный механизм, выполняющий определенные функции. Из субъединиц состоят все органы и ткани.
Система имеет цитоплазматическую мембрану, цитоплазму, ядро, также ряд органелл. Ядро разграничено с органеллами внутренней пленочкой. Все вместе обеспечивает жизнь тканям, а также позволяет осуществлять метаболизм.
Важную роль в функционировании играет цитоплазматическая плазма лемма или мембрана.
Само название наружная цитоплазматическая мембрана произошло от латин membrana или по другому кожица. Это разграничитель пространства между клеточными организмами.
Гипотезу строения выдвинули уже в 1935 г. В 1959 г. В. Робертсон пришел к выводу, что мембранные оболочки устроены по одному принципу.
Вследствие большого количества накопленной информации, полость приобрела жидко-мозаичную модель конструкции. Сейчас она считается признанной всеми. Именно наружная цитоплазматическая мембрана образует внешнюю оболочку единиц.
Строение
Итак, что такое плазма лемма?
Представляет собой тоненькую пленочку разграничивающую прокариоты с внутренней средой. Разглядеть ее можно только в микроскоп. В строение цитоплазматической мембраны входит би слой, который служит основой.
Би слой - это двойная прослойка, состоящая из белков и липидов. Также есть холестерол и гликолипиды, обладают амфипатричностью.
Что это значит?
Жировой организм имеет биполярную головку и гидрофильный хвостик. Первая обусловлена боязнью воды, а второй ее поглощением. Группа фосфатов имеет наружное направление от пленки, вторые направлены друг на друга.
Таким образом, происходит формирование биполярного липидного слоя. Липиды обладают высокой активностью, могут перемещаться в своем монослое, редко переходить в другие области.
Полимеры делятся на:
- наружные,
- интегральные,
- пронизывающие плазма лемму.
Первые находятся только на поверхностной части пазухи. Держатся за счет электростатики с биполярными головками липидных элементов. Удерживают питательные ферменты. Интегральные внутри, они встроены в саму структуру оболочки, соединения меняют свое местоположение за счет движения эукариот. Служат своеобразным конвейером, выстроены так, что по ним идут субстраты, продукты реакции. Белковые соединения пронизывающие макрополость имеют свойства образования пор для поступления питательных элементов в организм.
Ядро
В любой единице есть ядро, это ее основа. Цитоплазматическая мембрана также имеет органеллу, строение которого будет описано далее.
Ядерная структура включает пленку, сок, место сборки рибосом и хроматин. Оболочка разделена около ядерным пространством, оно окружено жидкостью.
Функции органеллы делятся на две основных:
- замыкание структуры в органелле,
- регулирование работы ядра и жидкого содержимого.
Ядро состоит из пор, каждая обусловливается наличием тяжелых поровых сочетаний. Их объем может говорить об активной двигательной способности эукариотов. Например, высокая активность незрелых содержит большее количество поровых областей. Ядерным соком служат белки.
Полимеры представляют соединение матрикса и нуклеоплазмы. Жидкость содержится внутри ядерной пленки, обеспечивает работоспособность генетического содержимого организмов. Белковый элемент выполняет защиту и прочность субъединиц.
В самом ядрышке созревают рибосомальные РНК. Сами гены РНК находятся на определенной области нескольких хромосом. В них происходит формирование маленьких организаторов. Внутри создаются сами ядрышки. Зоны в митозных хромосомах представлены сужениями, название вторичные перетяжки. При исследовании электроникой различают фазы фиброзного и грануляционного происхождения.
Развитие ядра
Другое обозначение фибриллярный, происходит из белковых и огромных полимеров-предыдущих версий р-РНК. В дальнейшем они образуют меньшие по размеру элементы зрелой р-РНК. Когда фибрилла созревает, она становится зернистой по структуре или рибонуклеопротеиновой гранулой.
Входящий в строение хроматин обладает окрашивающими свойствами. Присутствует в нуклеоплазме ядра, служит формой интерфазы жизнедеятельности хромосом. Состав хроматина, это нити ДНК и полимеры. Вместе они составляют комплекс нуклеопротеидов.
Гистоны выполняют функции организации пространства в структуре ДНК-молекулы. Дополнительно хромосомы включают органические вещества, ферменты, содержащие полисахариды, частицы металлов. Хроматин делится на:
- эухроматин,
- гетерохроматин.
Первый обусловлен низкой плотностью, поэтому считать генетические данные с таких эукариотов невозможно.
Второй вариант обладает компактными свойствами.
Структура
Сама конституция оболочки неоднородна. За счет постоянных движений на ней появляются наросты, выпуклости. Внутри это обусловлено движениями макромолекул и их выходом в другой слой.
Поступление самих веществ происходит 2 путями:
- фагоцитозом,
- пиноцитозом.
Фагоцитоз выражается во впячивании твердых частиц. Пиноцитозом называют выпуклости. Путем выпячивания, края областей смыкаются захватив жидкость между эукариотами.
Пиноцитоз осуществляет механизм проникновения соединений внутрь оболочки. Диаметр вакуоли составляет от 0,01 до 1,3 мкм. Далее вакуоль начинает погружение в цитоплазменный слой и от шнуровку. Связь между пузырьками играет роль транспортировки полезных частиц, расщеплении ферментов.
Цикл пищеварения
Весь круг пищеварительной функции разделяется на следующие этапы:
- попадание компонентов в организм,
- распад ферментов,
- попадание в цитоплазму,
- выведение.
Первая фаза подразумевает поступление веществ в тело человека. Далее они начинаются распадаться при помощи лизосом. Разделенные частички проникают в цитоплазменное поле. Непереваренные остатки просто выходят наружу естественным способом. Впоследствии пазуха становится плотной, начинается превращение в зернистые гранулы.
Функции мембраны
Итак, какие же функции она выполняет?
Главными будут:
- защитная,
- переносная,
- механическая,
- матричная,
- перенос энергии,
- рецепторная.
Защита выражается в барьере между субъединицей и внешней средой. Пленка служит регулятором обмена между ними. В результате последний может быть активным, либо пассивным. Происходит избирательность необходимых веществ.
При транспортной функции через оболочку передаются соединения от одного механизма к другому. Именно этот фактор влияет на доставку полезных соединений, выведение продуктов метаболизма и распада, секреторные компоненты. Вырабатываются градиенты ионного характера, благодаря чему идет поддержка ph и уровень концентрации ионов.
Последние две миссии относятся к вспомогательным. Работа на матричном уровне направлена на правильное расположение белковой цепочки внутри полости, их грамотное функционирование. За счет механической фазы клетка обеспечена в автономном режиме.
Перенос энергии происходит в результате фотосинтеза в зеленых пластидах, дыхательных процессов в клеточках внутри полости. В работе участвуют также белки. За счет нахождения в мембране белки снабжают макроклетку способностью воспринимать сигналы. Импульсы переходят от одной клетки-мишени к остальным.
К особым свойствам мембраны относят генерацию, осуществление биопотенциала, распознавание клеток, а то есть маркировка.
Цитоплазматическая мембрана, отделяющая цитоплазму от клеточной стенки, называется плазмалеммой (плазматической мембраной), а отделяющая ее от вакуоли - тонопластом (элементарной мембраной).
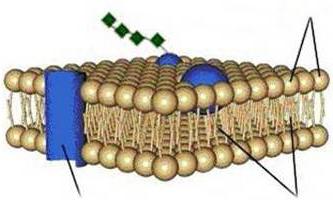
В настоящее время пользуются жидкостно-мозаичной моделью мембраны (рис. 1.9), в соответствии с которой мембрана состоит из бислоя липидных молекул (фосфолипидов) с гидрофильными головками и 2 гидрофобными хвостами, обращенными внутрь слоя. Помимо липидов в состав мембран входят белки.
Различают 3 типа мембранных белков, «плавающих» в билипидном слое: интегральные белки, пронизывающие всю толщу бислоя; полуинтегральные, пронизывающие бислой неполностью; перифери- ческие, прикрепляющиеся с внешней или внутренней стороны мембраны к другим мембранным белкам. Мембранные белки выполняют различные функции: одни из них являются ферментами, другие выполняют роль переносчиков специфических молекул через мембрану или образуют гидрофильные поры, через которые могут проходить полярные молекулы.
Одним из основных свойств клеточных мембран является их полупроницаемость: они пропускают воду, но не пропускают растворенные в ней вещества, т. е. обладают избирательной проницаемостью.
Рис. 1.9. Схема строения биологической мембраны:
А - внеклеточное пространство; Б - цитоплазма; 1 - бимолекулярный слой липидов; 2 - периферический белок; 3 - гидрофильная область интегрального белка; 4 - гидрофобная область интегрального белка; 5 - углеводная цепь
Транспорт через мембраны
В зависимости от затрат энергии транспорт веществ и ионов через мембрану делится на пассивный, не требующий затрат энергии, и активный, связанный с потреблением энергии. К пассивному транспорту относятся такие процессы, как диффузия, облегченная диффузия, осмос.
Диффузия - это процесс проникновения молекул через липидный бислой по градиенту концентраций (из области большей концентрации в область меньшей). Чем меньше молекула и чем более неполярная, тем быстрее она диффундирует через мембрану.
При облегченной диффузии прохождению вещества через мембрану помогает какой-либо транспортный белок. Таким образом, в клетку поступают различные полярные молекулы, такие, как сахара, амино- кислоты, нуклеотиды и др.
Осмос - это диффузия воды через полупроницаемые мембраны. Осмос вызывает передвижение воды из раствора с высоким водным потенциалом в раствор - с низким водным потенциалом.
Активный транспорт - это перенос молекул и ионов через мембрану, сопровождаемый энергетическими затратами. Активный транспорт идет против градиента концентрации и электрохимического градиента и использует энергию АТФ. В основе механизма активного транспорта веществ лежит работа протонного насоса (Н+ и К+) у растений и грибов, которые сохраняют внутри клетки высокую концентрацию К+ и низкую - Н+ (Na+ и К+ - у животных). Энергия, необходимая для работы этого насоса, поставляется в виде АТФ, синтезируемой в процессе клеточного дыхания.
Известна еще одна разновидность активного транспорта - эндо- и экзоцитоз. Это 2 активных процесса, с помощью которых различные молекулы транспортируются через мембрану в клетку (эндоцитоз ) либо из нее (экзоцитоз) .
При эндоцитозе вещества попадают в клетку в результате инвагинации (впячивания) плазматической мембраны. Образующиеся при этом пузырьки, или вакуоли, переносятся в цитоплазму вместе с заключенными в них веществами. Поглощение больших частиц, таких, как микроорганизмы или обломки клеток, называется фагоцитозом. В этом случае образуются крупные пузырьки, называемые вакуолями. Поглощение жидкостей (суспензий, коллоидных растворов) или растворенных веществ с помощью небольших пузырьков носит название пиноцитоз.
Обратный эндоцитозу процесс называется экзоцитозом. Многие вещества выводятся из клетки в специальных пузырьках или вакуолях. Примером может служить вывод из секреторных клеток их жидких секретов; другой пример - это участие пузырьков диктиосом в формировании клеточной оболочки.
ПРОИЗВОДНЫЕ ПРОТОПЛАСТА
Вакуоль
Вакуоль - это резервуар, ограниченный одинарной мембраной - тонопластом. В вакуоли содержится клеточный сок - концентрированный раствор различных веществ, таких, как минеральные соли, сахара, пигменты, органические кислоты, ферменты. В зрелых клетках вакуоли сливаются в одну, центральную.
В вакуолях хранятся различные вещества, в том числе конечные продукты обмена. От содержимого вакуоли в сильной степени зависят осмотические свойства клетки.
В связи с тем что вакуоли содержат крепкие растворы солей и других веществ, клетки растений постоянно осмотически поглощают воду и создают гидростатическое давление на клеточную стенку, называемое тургорным. Тургорному давлению противостоит равное ему по величине давление клеточной стенки, направленное внутрь клетки. Большинство растительных клеток существуют в гипотонической среде. Но если такую клетку поместить в гипертонический раствор, вода по законам осмоса начнет выходить из клетки (для выравнивания водного потенциала по обе стороны мембраны). Вакуоль при этом сократится в объеме, ее давление на протопласт уменьшится, и мембрана начнет отходить от клеточной стенки. Явление отхождения протопласта от клеточной стенки называется плазмолизом. В природных условиях такая потеря тургора в клетках приведет к увяданию растения, опусканию листьев и стеблей. Однако этот процесс обратим: если клетку поместить в воду (например, при поливе растения), возникает явление, обратное плазмолизу - деплазмолиз (см. рис. 1.10).
Рис. 1.10. Схема плазмолиза:
А - клетка в состоянии тургора (в изотоническом растворе); Б - начало плазмолиза (клетка, помещенная в 6% раствор КNО3); В - полный плазмолиз (клетка, помещенная в 10% раствор КNО3); 1 - хлоропласт; 2 - ядро; 3 - клеточная стенка; 4 - протопласт; 5 - центральная вакуоль
Включения
Клеточными включениями являются запасные и экскреторные вещества.
Запасные вещества (временно выключенные из обмена) и вместе с ними отбросы (экскреторные вещества) часто называют эргастическими веществами клетки. К запасным веществам относят запасные белки, жиры и углеводы. Эти вещества накапливаются в течение вегетационного периода в семенах, плодах, подземных органах растения и в сердцевине стебля.
Запасные вещества
Запасные белки, относящиеся к простым белкам - протеинам, чаще откладываются в семенах. Осаждающиеся белки в вакуолях образуют зерна округлой или эллиптической формы, называемые алейроновыми. Если алейроновые зерна не имеют заметной внутренней структуры и состоят из аморфного белка, их называют простыми. Если в алейроновых зернах среди аморфного белка встречаются кристаллоподобная структура (кристаллоид) и блестящие бесцветные тельца округлой формы (глобоиды), такие алейроновые зерна называют сложными (см. рис. 1.11). Аморфный белок алейронового зерна представлен гомогенным непрозрачным белком желтоватого цвета, набухающим в воде. Кристаллоиды имеют характерную для кристаллов ромбоэдрическую форму, но в отличие от истинных кристаллов составляющий их белок набухает в воде. Глобоиды состоят из кальциево-магниевой соли, содержат фосфор, нерастворимы в воде и не дают реакцию на белки.
Рис. 1.11. Сложные алейроновые зерна:
1 - поры в оболочке; 2 - глобоиды; 3 - аморфная белковая масса; 4 - кристаллоиды, погруженные в амфорную белковую массу
Запасные липиды обычно располагаются в гиалоплазме в виде капель и встречаются почти во всех растительных клетках. Это основной тип запасных питательных веществ большинства растений: наиболее богаты ими семена и плоды. Жиры (липиды) - наиболее калорийное запасное вещество. Реактивом на жироподобные вещества является судан III, окрашивающий их в оранжевый цвет.
Углеводы входят в состав каждой клетки в виде растворимых в воде сахаров (глюкозы, фруктозы, сахарозы) и нерастворимых в воде полисахаридов (целлюлозы, крахмала). В клетке углеводы играют роль источника энергии для реакций обмена веществ. Сахара, связываясь с другими биологическими веществами клетки, образуют гликозиды, а полисахариды с белками - гликопротеины. Состав углеводов растительной клетки значительно более разнообразен, чем у животных клеток, за счет разнообразного состава полисахаридов клеточной оболочки и сахаров клеточного сока вакуолей.
Главнейшим и наиболее распространенным запасным углеводом является полисахарид крахмал. Первичный ассимиляционный крахмал образуется в хлоропластах. Ночью, при прекращении фотосинтеза, крахмал гидролизуется до сахаров и транспортируется в запасающие ткани - клубни, луковицы, корневища. Там в особых типах лейкопластов - амилопластах - часть сахаров откладывается в виде зерен вторичного крахмала. Для крахмальных зерен характерна слоистость, что объясняется различным содержанием воды из-за неравномерного поступления крахмала в течение суток. В темных слоях воды больше, чем в светлых. Зерно с одним центром крахмалообразования в центре амилопласта называют простым концентрическим, если центр смещен - простым эксцентрическим. Зерно с несколькими крахмалообразующими центрами - сложное. У полусложных зерен новые слои откладываются вокруг нескольких крахмалообразующих центров, а затем формируются общие слои и покрывают крахмалообразующие центры (см. рис. 1.12). Реактивом на крахмал является раствор йода, дающий синее окрашивание.
Рис. 1.12. Крахмальные зерна картофеля (А):
1- простое зерно; 2 - полусложное; 3 - сложное; пшеницы (Б), овса (В)
Экскреторные вещества (продукты вторичного обмена)
К клеточным включениям относятся и экскреторные вещества, например кристаллы оксалата кальция (одиночные кристаллы, рафиды - игольчатые кристаллы, друзы - сростки кристаллов, кристалли- ческий песок - скопление множества мелких кристаллов) (см. рис. 1.13). Реже кристаллы состоят из карбоната кальция или кремнезема (цистолиты ; см. рис. 1.14). Цистолиты откладываются на клеточной стенке, вдающейся внутрь клетки в виде гроздьев винограда, и характерны, например, для представителей семейства крапивных, листьев фикуса.
В отличие от животных, выводящих избыток солей вместе с мочой, растения не имеют развитых органов выделения. Поэтому считается, что кристаллы оксалата кальция являются конечным продуктом метаболизма протопласта, образующимся как приспособление для выведения из обмена излишков кальция. Как правило, эти кристаллы накапливаются в органах, которые растение периодически сбрасывает (листья, кора).

Рис. 1.13. Формы кристаллов оксалата кальция в клетках:
1, 2 - рафида (недотрога; 1- вид сбоку, 2 - на поперечном срезе); 3 - друза (опунция); 4 - кристаллический песок (картофель); 5 - одиночный кристалл (ваниль)
Рис. 1.14. Цистолит (на поперечном срезе листа фикуса):
1 - кожица листа; 2 - цистолит
Эфирные масла скапливаются в листьях (мята, лаванда, шалфей), цветках (шиповник), плодах (цитрусовые) и семенах растений (укроп, анис). Эфирные масла не принимают участия в обмене веществ, но их широко используют в парфюмерии (розовое, жасминное масла), пищевой промышленности (анисовое, укропное масла), медицине (мятное, эвкалиптовое масла). Резервуарами для скопления эфирных масел могут быть желёзки (мята), лизигенные вместилища (цитрусовые), железистые волоски (герань).
Смолы - это комплексные соединения, образующиеся в процессе нормальной жизнедеятельности или в результате разрушения тканей. Они образуются эпителиальными клетками, выстилающими смоляные ходы, как побочный продукт обмена веществ, часто с эфирными маслами. Могут накапливаться в клеточном соке, цитоплазме в виде капель или во вместилищах. Они нерастворимы в воде, непроницаемы для микроорганизмов и благодаря своим антисептическим свойствам повышают сопротивляемость растений болезням. Применяются смолы в медицине, а также при изготовлении красок, лаков и смазочных масел. В современной промышленности заменяются синтетическими материалами.
Клеточная стенка
Жесткая клеточная стенка, окружающая клетку, состоит из целлюлозных микрофибрилл, погруженных в матрикс, в состав которого входят гемицеллюлозы и пектиновые вещества. Клеточная стенка обеспечивает механическую опору клетке, защиту протопласта и сохранение формы клетки. При этом клеточная стенка способна к растяжению. Являясь продуктом жизнедеятельности протопласта, стенка может расти только в контакте с ним. Через клеточную стенку происходит передвижение воды и минеральных солей, но для высокомолекулярных веществ она полностью или частично непроницаема. При отмирании протопласта стенка может продолжать выполнять функцию проведения воды. Наличие клеточной стенки более чем все другие признаки отличает растительные клетки от животных. Архитектуру клеточной стенки в значительной степени определяет целлюлоза. Мономером целлюлозы является глюкоза. Пучки молекулы целлюлозы формируют мицеллы, которые объединяются в более крупные пучки - микрофибриллы. Реактивом на целлюлозу является хлор-цинк-йод (Cl-Zn-I), дающий сине-фиолетовое окрашивание.
Целлюлозный каркас клеточной стенки заполнен нецеллюлозными молекулами матрикса. В состав матрикса входят полисахариды, называемые гемицеллюлозами; пектиновые вещества (пектин), очень близкие к гемицеллюлозам, и гликопротеиды. Пектиновые вещества, сливаясь между соседними клетками, образуют срединную пластинку, которая располагается между первичными оболочками соседних клеток. При растворении или разрушении срединной пластинки (что происходит в мякоти созревших плодов) возникает мацерация (от лат. maceratio - размягчение). Естественную мацерацию можно наблюдать у многих перезрелых плодов (арбуз, дыня, персик). Искусственную мацерацию (при обработке тканей щелочью или кислотой) используют для приготовления различных анатомических и гистологических препаратов.
Клеточная стенка в процессе жизнедеятельности может подвергаться различным видоизменениям - одревеснению, опробковению, ослизнению, кутинизации, минерализации (см. табл. l.4).
Таблица 1.4.
Похожая информация.